









































![Functional Anatomy of
Chordates -11
Nucleus
describe the origin and embryonic development of kidneys, gonads and their
associated ducts,
discuss the morphological and physiological adaptations of the urinogenital system
of vertebrates.
9.2 THE URINARY SYSTEM IN CHORDATES
9.2.1 Urinary System in Protochordates
The urinogenital system of protochordates such as the Cephalochordate [e.g.
Branchiostoma (amphioxus)] ahd Urochordate, (e.g. Herdmania) is very different both in
structure and origin from that of the vertebrates (Refer unit 1).
Cephalochordata
Despite the similarities to vertebrates in other aspects of its anatomy, the specialised
organs of excretion in the cephalochordate, Branchiostoma (amphioxus) show no
relationship to any part otthe vertebrate kidney or other known fluid regulating structure.
The organs of excretion in Branchiostoma are the protonephridia -which are
ectodermal in origin (unlike vertebrate kidneys which originate from the mesoderm).
Protonephridia(Fig, 9.1) are segmentally arranged sac-like tubes which lie in coelomic
spaces or atrium abbve the pharynx. Each protonephridium opens into the peribranchial
or atrial space surrdunding the gills so that its excretory products released into the atrium,
can be flushed away by the outgoing stream of water. Internally, within the coelomic sac
the protonephridium terminates blindly (not opening) into the coelom. (see also Fig.
9.17a in this unit)
Dorsopharyngeal coelom
./ Coelomic fluid
II Fig. 9:l: The protonephridium of Broncl~iostoma.
(Arrows show path of fluid flow)
i
The protonephridibl sac bears numerous, specialised tubular flame cells called the
solenocytes (Fig. 9.2.). Each solenocyte has a single flagellum projectrng downwards
into the protonephridial tube or canal. Constant beating of flagella probably forces fluid
into the protonephridia. The exact mechanism of functioning of solenocytes is still not
-Flagellum
clear. As the distql ends of solenocytes lie close to branchial (gills) blood vessels, the
solenocytes probably help to filter blood fluids from the branches of branchial blood
vessels. In the prdcess some fluid components such as ions are probably returned to the
body (Fig. 9.1).
Fig. 9.2: Enlarged view of a single
flame cell or solenocyte. Urochordata
In the urochordatg, Herdmania, neural gland is supposed to be the excretory organ (Fig.
9.3) and lies embedded in the mantle above the nerve ganglion or brain. The neural gland
consists of few central tubules from which arise peripheral tubules. The central tubules
open into a longitbdinal canal. The longitudinal canal opens by a ciliated funnel at the
base of dorsal tubercle. The excretory cells are called nephrocytes. They are present in
blood in order to kollect waste which are in the form of xanthene and urates. The waste
passes through th lumen of the neural gland and its duct and is finally discharged into
the pharynx. Neufa1 glands are endocrine in function as well, since they also control
oviposition, development and metamorphosis.](https://image.slidesharecdn.com/bsc-zoology-02sem-drvijay-comparativeanatomyofvertebrates-250829150549-91a61136/85/BSc-Zoology-02Sem-DrVijay-Comparative-anatomy-of-vertebrates-pdf-46-320.jpg)





































BSc-Zoology-02Sem-DrVijay-Comparative anatomy of vertebrates.pdf
- 1. WARM-UP: 1. WHAT ARE THE FUNCTIONS OF THE CIRCULATORY SYSTEM? 2. HOW DOES THE CIRCULATORY SYSTEM DEPEND ON THE RESPIRATORY SYSTEM? 3. DO YOU THINK ENDOTHERMS OR ECTOTHERMS NEED MORE EFFICIENT CIRCULATORY SYSTEMS? JUSTIFY YOUR ANSWER. Comparative Anatomy: Circulatory Systems in Vertebrates
- 2. Structures and Functions of the Circulatory System Functions Transports nutrients, oxygen, waste, hormones, and cells throughout the body. Helps stabilize body temperature Maintains the pH inside the body
- 3. Structures and Functions of the Circulatory System Functions Transports nutrients, oxygen, waste, hormones, and cells throughout the body. Helps stabilize body temperature Maintains the pH inside the body Structures Heart Blood Blood vessels: Veins, Arteries and Capillaries
- 4. Class Osteichthyes Internal Transport Closed circulatory system-blood is contained within blood vessels such as veins, arteries, and capillaries Two-chambered heart (atrium, ventricle)
- 5. Class Amphibia: Internal Transport Adults: Three chambered heart Improved heart to deliver more oxygen to walking muscles. Tadpoles have a two-chambered heart
- 6. Class Reptilia: Internal Transport Can be argued that they have a three or four chamber heart (both are correct) Crocodiles have a four chambered heart. All others have a partially devided ventricle
- 7. Four-chambered heart Class Aves: Internal Transport
- 9. Class Mammalia: Internal Transport 4-chambered heart
- 11. UNIT 10 NERVOUS SYSTEM AND SENSE ORGANS Structure 10.1 Introduction Objectives 10.2 Nervous Tissue in Vertebrates 10.3 Central Nervous System Cavities of the Brain and Spinal Cord The Spinal Cord The Brain 10.4 Peripheral Nervous System Splnal Nerves Cranial Nerves Autonomic Nerves 10.5 Brain -A Comparative Account Jawless Vertebrates Jawed Vertebrates 10.6 Sense Organs The Eye The Ear Olfactory Organs 10.7 Specialised Sensory Organs Lateral Line System in Fishes Pit Organs in Snakes Echolocation in Bats 10.8 Summary 10.9 Terminal Questions 10.10 Answers 1 0 . INTRODUCTION - - - - - - - - Each of us would have at some point in our life observed animals, how they move, catch prey or feed and respond to external and internal stimuli. All these activities take place because individual cells in the animal's body respond to certain stimuli and their responses are integrated in a meaningful co-ordinated manner. This co-ordination occurs in two forms, electrical through the nervous system and in chemical form through the endocrine system.ln this unit you will learn about the organisation of the vertebrate nervous system while the other integrating system will be dealt with in Unit 12. You have studied in LSE-05, and LSE-09 how specialised cells of the metazoan body the neurons, get organised into a nervous system and that these neurons operate on the same principles through out the animal kingdom. You would recall that the function of the nervous system is to receive stimuli or sensory information and to send irnp~llses from one part of the body to another. In this manner it regulates an animal's activities by integrating incoming sensory information with stored information, the result of past experience and then translating past and present information into action through effectors. Nervous tissue is also the seat of all conscious experience. In this unit you will learn briefly about the structure of a vertebrate nerve cell which is the functional and structural unit of the nervous system. We will discuss the organisation of the vertebrate nervous system and the brain in relation to function in different vertebrate groups. To be able to respond to the external and internal stimuli , animals possess sensory receptors which may be wide spread in the body or may be in the form of specialised organs the sense organs. These translate environmental energies into electrical impulses that are transmitted to the nervous system. We describe the three basic types of sense organs, the optic, auditory and olfactory organs. Vertebrates differ in their ability to perceive stimuli, hence some vertebrate groups have specialised sense organs that have originated to suit their special mode of life. They too would be discussed briefly to emphasise how these organs help the animal to respond to its external environment.
- 12. ~unctional Anatomy of Chordates - 11 Objectives After studyingthis unit you should be able to: describe the central, peripheral and autonomic nervous system of vertebrates, give a comparative account of brain in vertebrates, correlate the evolution of brain structures with their function in vertebrates, illustratethe structureof eye, internal ear and olfactory organs in vertebrates, describe specialisedsensdry organs. - 10.2 NERVOUS TISSUE IN VERTEBRATES You have learnt in Block - 3 of the DevelopmentalBiology Course (LSE-06) that all nervous tissue is ectodermal in origin. In vertebrates during embryonic development the flattened layer of ectoderm along the mid dorsal side of gastrula becomes thickened and is known as neural or medullary plate which gives rise to the neural tube and neural crest. The neural tube is the forerunner of the brain and spinal cord, and some of the neural crest cells migrate away from the neural tube to give hse to the bodies of neurons that lie outside the brain and spinal cord. You would recall f r m earlier courses LSE-05 and LSE-09 the structure of a typical neuron which consists of a cell body and several processes arising from it - the dendrites, usually numerous and highly branched and the single long process the axon with branches- the terminal arboration at its end (Fig. lO.l).Collateral branches may be given off from the axon but often these are lacking. The axon terminal may make close contact (synapse) with the dendrites of another neuron and neurotransmitters are released at the axon terminals that conduct the information in the form of impulses across the synapse to the other neuron. This is normally unidirectional. A single neuron may have contact with thousands of other neurons that transmit information along their axons aild receive information over dendrites. eurolnusc~llar sy::apse Muscle Cell (b). , Dendrites Fig. 10.1 :Typical neuronsin vertebrates. a ) Somatic sensory. b) Somatic motor. c ) Granule cell (cerebellar cortex ).d) Purkinje cell (cerebellar cortex). Though the basic componentsof the nervous system is the neuron, another kind of tissue the neuroglia (nerve glue) are interspersed among the nervous elements and provide support and slome degree of protection. These do not conduct signals nor emit neurotransmitters. The two principal types of neuroglia are 1) macroglia of ectddermal origin.
- 13. 2) microglia of mesodermal origin. Nervous System and Senlse Organs One kind of macroglia are the oligodendrite cells. These extend processes that wrap around axons. This wrapping or sheath is composed of myelin a substance rich in fats and proteins. Myelin sheaths are present generally in axons of only vertebrates. Axons of neurons outside the brain and spinal cord are also coated by ribbon like cells. These are Schwann cells and are similar to oligodendrites in that they also produce myelin that serves as insulating material which in the manner of coating on an electric wire, prevents loss of energy of the nerve impulse during its passage along the axon. Presence of myelin sheath also helps in fast conduction of nerve impulse as the fibres with thick covering of myelin conduct at the greatest speed. This myelin sheath is interrupted at regular intervals by circular constrictions forming the nodes of Ranvier. Amongst the vertebrates, myelin sheaths are absent in cyclostomes. Another kind of microglial cells are the astrocytes which are the largest and the most abundant. They make contact with other nervous tissue and maintain normal nervous tissue physiology. They also play a role in brain development repair and healing and also maintaining the blood-brain barrier. A collection of nerve cell bodies is known as a ganglion. Groups of nerve cell bodies and their dendrites and the proximal unmyelinated portion of the axons have a greyish appearance, they form the grey matter. The brain and spinal cord are chiefly composed of grey matter. On the other hand, white matter is composed of bundles of myelinated fibres. Such bundles are known as nerve tracts in the brain and spinal cord and nerve in the rest of the body. White and grey matter are sometimes intermingled. Such an arrangement is known as reticular formation. The vertebrate nervous system has two main division. I) The central nervous system (CNS) which consists of the brain and spinal cord. 2) Peripheral nervous system (PNS) consisting of the cranial nerves arising from the brain and the nerves and ganglia arising from the spinal cord. Part of the peripheral nervous system is composed of autonomic nerves which are distributed to those parts of the body that are under involuntary control. Let us now consider the central nervous system. But before you move onto the next Forebrain I Ii~~dbr;~i~i section try the SAQ given below. Midbrain A SAQ 1 Correct the following statements suitably: a) Spinal cord and brain are formed of white matter. b) Myelin sheaths are secreted by astrocytes. c) Myelin sheaths are found only in the axons of the brain paleopallium Eye lo lecluln I I.;II and I;l~erolline to cerebellum and medulla d) Bundles of axon in the brain form the reticular formation. Lateral venlricle / 10.3 CENTRAL NERVOUS SYSTEM 111 Venlricle L The central nervous system is composed of the brain, which lies within the cranial cavity of the skull and the spinal cord lying within the neural canal formed by the neural arches of the vertebrae. With the differentiation of the neural tube of the embryo into the brain IV ventricle and spinal cord its original cavity becomes modified to form the fluid filled ventricles El+ Foramenof nionro that are connected spaces located within the centre of the brain and the narrow central canal of the spinal cord. (b) Fig. 10.2 :a) Diagrammatic view of The anterior end of the neural canal can be recognised into three embryonic regions, the major subdivisions of prosencephalon, mesencephalon and rhombencephalon. These form the forebrain, the primitive vertebrate midbrain and hindbrain in the adult (Fig. 10.2). brain and their connections to the sense 10.3.1 Cavities of the Brain and Spinal Cord The anterior end of the prosencepha1,ongives rise to the telencephalon that ultimately forms the two cerebral hemispheres in the higher forms; with the development of the cerebral hemispheres the cavities extending into them become lateral ventricles or organs. b) Ventricles of the brain.
- 14. Functional Anatomy of Chordates - I[ ventricle I and I1 (see Fig. 10.2 b.). The remainder of the prosencephalon forms the diencephalon and its cavity is known as the third ventricle. Ventricles I, I1 communicate with ventricle 111by means of interventricular foramen or foramen of Monro. In higher vertebrates the IIIrd ventricle communicates with mesencephalon by cerebral aqueduct a narrow canal and posteriorly the cerebral aqueduct leads into the fourth ventricle in the rhombencephalon. The portion of the fourth ventricle within medulla oblongata is refe~ed to as myelocoele which is continuous with the canal of the spinal cord. The cavitlies of the brain and spinal cord are filled with lymph like cerebrospinal fluid. 10.3.2 The Spinal Cord The portion of the neural tube which forms the spinal cord undergoes considerably less modification than that forming the brain. It generally assumes the shape of a more or less cylindrical, but slightly flattened tube. It widens at the anterior end, where it is continuous with the medulla oblongata. The posterior end usually tapers down to a fine thread the filum terminale. In cyclostomes and fisheg the spinal cord is fairly uniform in diameter but in most tetrapods two prominent swellings or enlargements are seen where the nerves going to the limbs arise. In the anterior part the cervical enlargement is the region where the large nerves supplying the forelimbs arise and the lumbar enlargement, near the posterior end of the cord where the nerves supplying the hindlimbs originate. In limbless forms such as snakes neither enlargement is seen. Grey and white matter of the cord In cross section, the spinal cord is seen to be composed of grey and white matter. The grey matter is almost completely surrounded by the white matter. The grey matter in amniotes is arranged in the shape of the letter 'H' (Fig. 10.3). The portions corresponding to the upper bars of the 'HI' extend dorsally and are known as dorsal columns and the lower bars, the ventral ~columns. The connecting bar in which the central canal lies forms the dorsal and ventral grey commissures, above and below respectively (Fig. 10.3 a). ,Dorsal septum Dorsal column Lateral funiculus Ventral col~unn , Somatic sensory colu~nn Visceral sensory colu~nn isccral motor column Somitic motor colllmn Fig. 10.3 :a) A cross section ofspinal cord of cat, b) Relative positions of the four columns of grey matter in each side of spihal cord.
- 15. The cell bodies in the dorsal columns are, for the most part, those of association neurons. Their dendrites form synapses with the axons of sensory or afferent nerve fibres which enter the spinal cord via the dorsal roots of spinal nerves . The axons of the association neurons form synapses with the dendrites of motor or efferent neurons, the cell bodies of which are located in the ventral columns. The somatic sensory fibres carry impulses from somatic tissue to central nervous system. They form synapses with cells in the upper portion of dorsal columns, while visceral sensory fibres form synapses with cells in lower portion ofthe dorsal column (Fig. 10.3 b). T k cell bodies of somatic motor neurons and fibres are located in the lower portions of the ventral column (Fig. 10.3 b), whereas, the cell bodies of visceral motor neurons and fibres have their origin in the upper and lateral portions of the ventral columns. The white matter of the cord is arranged in longitudinal columns called funiculi which lie outside the grey matter (Fig. 10.3 a). The funiculi are divided into fibre tracts, composed of medullated fibres, carrying impulses up and down the cord and to and from the brain. Lateral funiculi lie between dorsal and ventral columns, the dorsal funiculus lies between the dorsal septum and dorsal column and a ventral funiculus is located between the ventral fissure and ventral column of grey matter .The two ventral funiculi are connected through the ventral commissure. The dorsal funiculi carry the sensory nerve impulses up the cord and to the brain, while those in the ventral funiculi are primarily motor, carrying impulses down the cord and from the brain. The lateral funiculi carry both sensory and motor fibres. Lower vertebrates do not have such an elaborate arrangement ofcolumns and funiculi. In amphioxus there is no clear cut distinction between white and grey matter as medullated fibres have not appeared. In cyclostomes there is yet no sharp delineation between grey and white matter in the spinal cord. SAQ 2 a) Match the following Telencephalon Diencephalon Rhombencephalon Medulla oblongala Myelocoel Fourth ventricle Lateral ventricles Third ventricle b) Fill in the blanks with appropriate words from the text. i) The dorsal column of the spinal cord has cell bodies of ..................... neurons. ii) Sensory nerve fibres enter the spinal cord via the ..................roots. iii) Ventral columns of the spinal cord contain the cell bodies o f ................... neurons. iv) ..................................... carry messages upto the brain while ........................................ carry messages down the cord from the brain 10.3.3 The Brain The chordate brain is basically an enlargement of the anterior end of the neural tube. In primitive condition the cell bodies of the neurons comprising the central nervous system are aggregated around the central canal of the neural tube. Although this arrangement persists in the spinal cord, in the brain region, migration of cells to the peripheral areas occurs. In amphioxus the brain is seen in its simplest form, as a cerebral vesicle. As the vertebrates evolved their brain grew larger and more complex, and the various parts of the brain developed to suit the specialised demands of their particular environment. For example, cavefish, that live in permanently dark subterranean environments have reduced eyes. Correspondingly, the part of the brain which normally receives visual input is reduced as well. In salmon, on the other hand the same portion is enlarged as visual information constitutes a large part of the sensory input to the brain. Nervous System and Sense Organs All functions in the body can be termed as either somatic or visceral. Somatic functions are those carried out by the skin and its derivatives. voluntary muscles and skeletal structures. Visceral functions are performed by other organ system of the body i.e. digestive, respiratory etc.
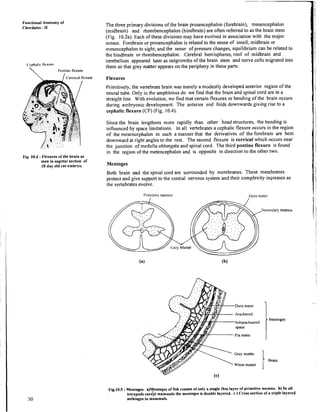
- 16. Functional Anatomy of Chordates - I1 The three primary divisions of the brain prosencephalon (forebrain), mesencephalon (midbrain) and rhombencephalon (hindbrain) are often referred to as the brain stem (Fig. 10.2a). Each of these divisions may have evolved in association with the major senses. Forebrain or prosencephalon is related to the sense of smell; midbrain or tnesencephalon to sight; and the sense of pressure changes, equilibrium can be related to the hindbrain or rhombencephalon. Cerebral hemispheres, roof of midbrain and cerebellum appeared later as outgrowths of the brain stem and nerve cells migrated into C'cplial~c Ilexvrr them so that grey matter appears on the periphery in these parts. I'ontinc flexure I Fig. 10.4 :Flexures of the brain as seen in sagittal section of 18 day old rat embryo. Flexures Primitively, the vertebrate brain was merely a modestly developed anterior region of the neural tube. Only in the amphioxus do we find that the brain and spinal cord are in a straight line. With evolution, we find that certain flexures or bending of the brain occurs during embryonic development. The anterior end folds downwards giving rise to a cephalic flexure (CF) (Fig. 10.4). Since the brain lengthens more rapidly than other 'head structures, the bending is influenced by space limitations. In all vertebrates a cephalic flexure occurs in the region of the mesencephalon in such a manner that the derivatives of the forebrain are bent downward at right angles to the rest. The second flexure is cervical which occurs near the junction of medulla oblongata and spinal cord. The third pontine flexure is found in the region of the metencephalon and is opposite in direction to the other two. Meninges Both brain and the spinal cord are surrounded by membranes. These membranes protect and give support to the central nervous system and their complexity increases as the vertebrates evolve. I'ria~itivcmeninx I>ura mater # v Arachnoid , ) Y : ; ~ a c h a n o i d 7 .St Pia mater Grey mattei White matter IMeninges Fig.1O.S :Meninges. ajyeninges of fish consist of only a single thin layer of primitive meninx. b) In all tetrapods exkept mammals the meninges is double layered. c ) Cross section of a triple layered meninges in mammals.
- 17. Cartilage and bone are covered with a tough vascular membrane, perichondirum, that lines the cavities in which the brain and spinal cord lie. In cyclostomes and fishes (Fig. 10.5a) a single membrane, meninx pri~nitiva forms a close union with brain and spinal cord. With the adoption of terrestrial life the meninges doubled. In amphibians, reptiles and birds (Fig. 10.5b), instead of a single meninx, an inner pia-arachnoid layer and outer dura mater is observed. The cavity between these two layers is filled with cerebrospinal fluid that protects the brain and spinal cord from shocks during terrestrial !ocomotion. In mammals ( Fig. 10.5 c ) the tough dura mater persists and pia- arachnoid membrane differentiates into two layers, an inner pia mater and an outer arachnoid membrane. The pia mater contains blood vessels that supply the underlying nervous tissue. A subarachnoid space filled with cerebrospinal fluid makes its appearance between the two. In the brain region the cranial dura mater fuses with the endorachis and the epidural space and thus disappears. The cerebrospinal fluid, present in the ventricles of the brain, circulates slowly through the various cavities and spaces between the membranes. Grey and white matter of the brain The grey matter of the brain like the spinal cord consists of nerve cell bodies with their dendrites and proximal portions of their axons. They are usually together in the form of nuclei. The white matter consists of tracts of myelinated fibres connecting various parts df the brain and of ascending and descending fibres carrying impulses to and from the spinal cord. Let us now consider the structure of the vertebrate brain as it has evolved from the primitive chordates to the advanced mammals. It would be better if you would read this description while referring to Figure 10.6 closely. I'rosmcephalon Mese~lcephalon Rhombrncrphalon -1- C'crebralhemisphere A Hypothalamusyentrd thalamus Fig. 10.6: Strttcturc of gettrralised brain in vertebrates. A) Prosencrphalott cl)t~sists of (1) tclcncepltalon and (2)diettccphalons. Rltombencepltalon consists of (3) ~~~etcttccpltalon and (4) myelerrccphalon, (B) Iatcral view of genet-alised brain showing ventriclrs. Hindbrain consists of the myelencephalon and metencephalon (Fig. 10.6 A). Nervous System and Sense Organs The cerebrospinal fluid is derived from blood and returns to it while circulating over the nervous tissue and in the ventricle of the brain. It is however, devoid of all red blood cells as well as other large formed elements. When a person is injured and trauma to the CNS is suspected, the cerebrospinal fluid sample is taken. If it contains red blood cells then the brain or spinal cord may be damaged. Myelencephaloil is the posterior most portion of the hindbrain and merges with the spinal cord. It forms the medulla oblongata. It is also referred to as the oldest part of the brain as it is well developed in all vertebrates even though other portions may be rudimentary. The general structure of medulla is like the spinal cord except that the central canal enlarges (as the 4th ventricle) and a thin highly vascular roof known as posterior choroid
- 18. Functional Anatomy of Cl~ordates - I1 plexus forms dorsal to the central canal. The choroid plexi of the brain produce the cerebrospinal fluid and control its composition. The medulla conltains important nerve centres or nuclei which control vital physiological fwhctions that are involuntary like heartbeat, respiration and metabolism. It also houses the primary nuclei of cranial nerves. Damage to the medulla can be life threatening. The dorsal anterior portion of the medulla contains-nuclei associated with the nerves from lateral line system and inner ear. In terrestrial vertebrates these nuclei are associated with equilibrium and auditory functions of the ear. The medulla also serves as a route for descending and ascending pathways that run from and to the higher brain centres. Metencephalon is the anterior part of the hindbrain, the dorsal part of which becomes elevated and thickened to form the cerebellum. The cerebellum is highly developed in animals that are active and whose balance and precise motor movements are well developed whether in water, air or on land. The function of the cerebellum is to monitor and modify motor outputs but it does not initiate them. It operates at an involuntary level and maintains equilibrium. Information regarding touch, vision, hearing, proprioreception (related to limb position, joint angle, state of muscle contraction) and motor input from higher centres of the brain are processed in the cerebellum. For an organism to tly, jump swim in a three dimensional world in space in relation to gravity, the cerebellum is involved in maintaining its positional equilibrium. Another function of cerebellum is refinement of motor action, removal of cerebellum will still allow the organism to move but the movement would be uncoordinated. The size of the cerebellum is proportional to its role. In fishes and amphibians (salamanders) the cerebellum is small and simple, as their locomotion is simple and mainly co-ordinated by the spinal reflexes. In advanced tetrapods such as mammals and birds there are so many cells in the cortex that it is folded. The increase in cells also increases the number of fibres and these fibres form bulging masses. The ventral side of metencephalon of mammals and some birds is composed of a prominent mass of transverse nerve fibres known as pons. Motor fibres from cerabral cortex pass via the pons to the cerebellum. Midbrain (Figs. 10.6A and B) or mesencephalon is marked off from the hindbrain very early in development by a conspicuous constriction, the isthmus. The floor and walls of the mesencephalon are thick and composed of fibre tracts, cerebral peduncles connecting forebrain and hindbrain. The roof consists of a thick layer of grey matter, the optic:tectum. The central canal of the midbrain is of a relatively small diameter forming a canal the cerebral aqueduct between the hindbrain and midbrain. In lower vertebrates two optic lobes arise from the roof. The optic lobes in lower vertebrates serve as centres for the visual sense. In higher forms however the optic lobes are practically solid and the roof is known as tectum. In fishes and amphibians the midbrain is often the most prominent region of the brain as all visual information is received here directly from the eyes.The anterior region of tectum in snakes and mammals is specialised into superior colliculi which serves to integrate visual inputs and posterior region is known as inferior colliculi, which integrates auditory inputs. Thus, visual information in all vertebrates reaches the forebrain via the tectum. In mammals however, due to development of cerebral hemispheres the superior colliculi are less important as visual centres. Forebrain consists of the diencephalon and the telencephalon. Diencephalon is the initial portion of the forebrain and regulates many bodily functions. Jt contains the expanded neural canal or third ventricle (Figs. 10.6 A and B) with a thin dorsal roof, the epithalamus, the walls form the thalamus and the floor forms the hypothalamus. Each is thickened by the presence of a proliferation of neurons. The epithalamus is made up o f a parietal body or parapineal body and pineal gland in lowdr vertebrates. The parietal body is absent from most higher vertebrates, only the pineal body or gland is present in all. The pineal body affects skin pigmentation in lower vertebrates and probably affects photoperiod. In higher vertebrates it is important in controlling biological rhythms. The thalamus contains a number of cell clusters that are important in co-ordinating sensory impulses from all parts of the body, except olfactory impulses that go directly
- 19. to the cerebral cortex. The thalamus is actually a relay centre for all sensory information going to the cerebral cortex. The anterior part of the roof of the diencephalon forms the anterior choroid plexus. The hypothalamus is themost ventral portion of diencephalon. This includes the posterior pituitary (neurohypophysis). Hypothalamic nuclei are involved in maintaining the body's internal homeostasis. They regulate appetite, sexual activity, body temperature, water balance and alertness and some aspects of emotional behaviour. Hypothala~nus stimulates the pituitary gland to regulate many homeostatic functions. Telencephalon or cerebrum is the terminal portion of the forebrain. It shows the greatest difference in degree of development among vertebrates. In primitive vertebrates the forebrain is mainly concerned with integrating sensory inputs from nasal olfactory sensors which are important in complex aspects of behaviour. From both sides of telencephalon arise the cerebral hemispheres that are associated with paired olfactory bulbs. The neural canal extends into the lateral ventricles of the cerebral hemispheres. (In amphioxus the neural canal does not branch as there are no cerebral hemispheres nor olfactory bulbs). The cerebral hemispheres enlarge to an increasing degree as the vertebrate scale is ascended and in the highest forms the cerebral hemispheres cover over the greatest part of the remainder of the brain. The seat of consciousness lies in the cerebral hemispheres. The nerve centres controlling the activities which characterise the highly developed psychic life of man, such as intelligence, thought and sensation are located in this region. At the anterior end of each hemisphere is an outgrowth called the olfactory lobe (Fig. l0.6B). The olfactory lobe may come in contact with the nasal apparatus. In the lowest living vertebrates, the hemispheres are divided into an anterior and posterior olfactory lobe, concerned mainly with receiving olfactory impulses that are relayed to the diencephalon. In all vertebrates the floor of each hemisphere differentiates into a thickened corpus striatum. The grey regions of corpus striatum are often referred to as basal nuclei . The remainder of the hemisphere consists of a pallium which form a roof over the Lateral ventricles . It is the pallium that has become so highly developed and modified in the evolution of the higher groups of vertebrates. In fishes the pallium is thin-walled and the grey matter is present only on its inner walls adjacent to the ventricle and the telencephelon serves mainly as an olfactory centre. As the vertebrate scale is ascended ,there is an increasing tendency for nerve cells from the inner grey layer to migrate out into peripheral area. The first real change is seen in reptiles. The cerebral hemispheres enlarge in size and extend backwards to cover partially the diencephalon and increased grey matter migrates to the periphery. A new area the neopallium appears in the reptiles and this is what forms the large cerebral hemispheres in mammals. In crocodile for the first time nerve cells migrate to the outer surface in the neopallium and form the true cerebral cortex. In ~nammals the neopallium enlarges enormously and the grey cell bodies form a layer of grey matter which even in humans is only a few centimetres thick. In all vertebrates below mammals, the cerebral hemispheres though large, are smooth. In many mammals the surface becomes convoluted or folded. The ridges are called gyri and depressions sulci. These convolutions increase the surface area and total amount of grey matter. Larger mammals have more convolutions though these are not necessarily connected to intelligence! Decussation Commissures serve to connect similar regions in the left and right sides of the central nervous system, and make bilateral integration possible. There are also fibre tracts in the brain which in their course, cross over, or decussate to the opposite side. Injury to one side of the brain often results in paralysis of muscles of the opposite side. Nervous System and Sense Organs In ncurophysiologya nucleus is a small clusteror aggregate of nerve cell bodies within the central nervou system.
- 20. Functional Anatoniy of 1 Chordates - I1 SAQ 3 a) Match the following correctly. 1 Metencephalon Medulla oblongata Mesencephalon Cerebellum Diencephalon Optic tectum Telencephalon Epithalamus, hypothalamus thalamus Myllencephalw. Cerebrum b) Fill in the blanks: ........................ i) In fishes the covers the brain and spinal cord. .................. ii) Reptiles and birds have a double membrane made up of and ....................to protect the brain. iii) In mammals the layers of the meninges are called ..................... ...........................and ...................... 10.4 PERIPHERAL NERVOUS SYSTEM The nerves and ganglia which form connections with the central nervous system and which are distributed to all parts of the body, comprise the peripheral nervous system or PNS. The autonomic portion of the peripheral nervous system is composed 'of those nerve fibres distributed to structures under involuntary control. Connection with the central nervous system is mediated via spinal and cranial nerves. 10.4.1 Spinal Newes Each spinal nerve connects to the spinal cord by means of two roots, dorsal and ventral. Dorsal roots originate from neural crests. A band of neural-crest cells is present on either side of the spinal cord and extends in a longitudinal direction in the embryo. Dorsal root Dorsal root ganglioll ~~mbathetic chain Ramus communicans Fig. 10.7 :Spinal cord and spinal nerve anatomy. a) Sensory ahd motor routes in the spinal nerve. b) Dorsal and ventral roots connect the spinal nerve to the spinal cord. Spinal nerve joins the autonomic chain ganglion through a communicrting ralnus.
- 21. At metameric intervals in each band enlargements occur and the parts of the band between enlargements gradually disappear. In these thickenings, each neuroblast sends out two processes: 1) an axon which grows towards the spinal cord and enters in the region of the dorsal columns and 2) a dendrite, which grows peripherally to the skin, voluntary muscles, skeleton, or some visceral structures. These thickenings form the dorsal root ganglion ( Fig. 10.7). The ventral roots arise from neurons in the grey matter of the ventral column of the spinal cord. The sensory nerve impulses travel towards the spinal cord via the dorsal roots and are spoken of as afferent fibres. The sensory fibres are said to be either somatic or visceral. Somatic sensory fibres are those coming from the skin and its derivatives, voluntary muscles and skeletal structures. They form synapses with cells in the somatic sensory column of grey matter. Visceral sensory fibres from visceral structures terminate in the visceral sensory column of the grey matter of the cord ( see Fig. 10.3 b again ). Cell bodies of the neurons making up the ventral roots lie in the grey matter of ventral columns of the spinal cord. In motor fibres, impulses are conveyed away from the spinal cord, hence they are referred to as efferent fibres. Somatic motor fibres arise in the somatic motor column of grey matter in the spinal cord and are distributed to somatic structures .Visceral motor fibres pass to autonomic ganglia, where they form synapses with motor autonomic neurons ( Fig. 10.7 b). The dorsal roots are strictly sensory in the amniotes. In amphioxus and lamprey dorsal roots are composed of both sensory and motor fibres. In fishesa&hibians visceral efferent fibres pass through both dorsal and ventral roots. Near the area where dorsal and ventral roots unite to form spinal nerves, three rami are usually given off. These include 1) a dorsal ramus supplying skin and muscles of dorsal part of the body, 2) a ventral ramus distributed to the ventral and lateral regions, and 3) a visceral ramus that forms connections with one of the chain ganglia of the peripheral autonomic nervous system (Fig.lO.7 b). Both dorsal and ventral rami are composed of somatic sensory and somatic motor fibres. A typical visceral ramus consists of a white ramus and a grey rarnus. The white ramus carries medullated visceral sensory and medullated preganglionic motor fibres. The grey visceral ramus carries only non-medullated postganglionic autonomic motor fibres. The fibres of grey ramus join the spinal nerve and travel out either via dorsal or ventral rarni where they supply structures under involuntary control such as blood vessels, muscles and glands of the skin. 10.4.2 Cranial Newes The peripheral nerves which form connections with the brain are called cranial nerves. There are 10 pairs of cranial nerves in anamniotes and 12 in amniotes. All but the first four are joined to the medulla oblongata. Some are entirely sensory, composed of afferent fibres alone; others are purely motor. Still others are mixed nerves, consisting of both motor and sensory fibres. The nature and distribution ofdifferent cranial nerves is given Table 10.I. Table 10.1: Cranial nerves, their nature and distribution. lachrymal gland, nose and forehead skin, Nervous Systcm and Sense Organs Early human anatomists assigned them numbers in an anterior posterior sequence. This system has now proved to be artificial and superficial but continues because of convenience and familiarly. 1.ater in 1894 a new cranial nerve was discovered which was numbered 0 to preserve the terminology of 1-10, or 1-12.
- 22. Functional Anatomy of Chordates - 11 I Maxillary Somatic Sensory rFrom upper jaw, upper lip, lower eyelid, teeth of upper jaw. Froin lower lip, teeth of lowerjaw. skin of temporal region, external ear. lower part of VI VII Vlll IX Sympathetic chain ganglia 111 Mandibular Abducens Facial X XI XI1 Sympathetic supply to sk'ili U Muscous membranes and blood vessel.;' Auditory Glossopharyngeal Fig.1O.R :Autonomic nervous system in humans. ~ e j l s ~ m ~ a t h e t i c division postganglionic fibres shown in dotted lines. Right parasympathetic division. Mixed Somatic Motor Mixed Vagus Spinal accessory Hypoglossal the +aceto i&scles used in chewing: To eye muscle, nictitating membrane. To muscles of face, scalp. external ear, lacrimal trland. lnucous membrane of nose Special soinatic Sensory Mixed From inner ear. From posterior region of tongue and taste buds. Mixed Visceral Motor Somatic Motor To lower jaw and throat, larynx and salivary glands. To pllarynx, larynx. (Considered as posterior branch of vagus) Muscles of tongue and muscles below the tongue in the lowerjaw
- 23. 10.4.3 Autonomic Newes The autonomic portion of the peripheral nervous system is composed of both sensory and motor fibres. Autonomic sensory fibres monitor the internal environmentof the organism, that is, blood pressure, oxygen and carbon dioxide tension, core and skin temperature and activity of the viscera, while the autonomic motor fibres send i~npulses to smooth mu'qcles and glands in various parts of the body. Autonomic nervous system regulates the functionsof structures which are under involuntary control.The proper functioningof this part of the nervous system is necessary for regulating such activitiesas rate of heartbeat, respiratory movements,composition of body fluids, constancy of temperature, secretion of various glands, peristalsis, and other vital processes and it is controlled by hypothalamic centres. Nervous System and Sense Organs Even though autonomicsystem is not under voluntary control, conscious centres also can atTect visceral activity controlled by the autonomic nervous system. For example through practice of meditation or through deliberated effort it is possible to atTect the heart beat or release of sweat. The ancie~~t yogis mastered the art. The system is composed of preganglionic and postganglionic fibres and a number of ganglia which serve as relay centres (Fig. 10.7). The cell bodies of the preganglionicneurons are located in the visceral motor column of grey matter in the central nervous system. Their medullated fibres pass to outlying ganglia. Postganglionicneurons have grey nonmedullated axons. The cell bodies of these neurons, lie in outlying ganglia which are often located at some distance from the central nervous system. It is here, that the preganglionic fibres form synapseswith the dendrites of postganglionicneurons . The autonomic portion of the peripheral nervous system is divided into two main parts, the sympatheticand parasympathetic system. The essential parts of both the systems are summarised in Figure 10.8. Both these systems work antagonistically.The sympathetic system functions.to strengthen the body reactions against adverse condition, it calls for expenditureof energy. The parasympatheticsystem is concerned with restoring and conservingenergy. In mammals, almost every visceral organ has sympatheticand parasympathetic innervation except the adrenal gland, peripheral blood vessels and sweat glands; all of which receive only sympathetic innervation.Cessation of sympatheticstimulation allows these organs to return to resting state. Post ganglionic axons of the sympathetic system except those going to uterus and sweat glands secrete norepinephrine or epinephrine (also known as noradrenalineor adrenaline). Thq postganglionic axons of the parasympatheticsystem release the neurotransmitter acetylcholine. Acetylcholine is also released between pre- and post ganglionic fibres in both these divisions of the autonomic system (Fig.lO.9). The sympatheticand parasympatheticfunctional componentsare clear in mammals, however, in other vertebrates their comparativeanatomy is not well understood. Sympathetic cord ~cet;lcholine Fig. 10.9 :Neurotransmitters of the autonomic system. Epinephrine and acetylcholine are released rt the post ganglionic nerve eqdings of the sympathetic and parasympathetic circuits respectively. This is the basis of their'antrgonistic functions. Sympathetic nervous system The visceral motor neurons that participate in sympathaticactivity depart from the thoracic and lumbar regions of the spinal cord, hence it is also known as 'thoracolumbar outflow'.
- 24. FunctionalAnatomy of Chordates-I1 On either side of the ventral part of the vertebral column lies a long sympathetic trunk, extending from foramen magnum to the coccyx. At fairly regular intervals, each sympathetic trunk lbears enlargements known as chain ganglia (Fig.10.8). The chain ganglia are numbered according to the vertebrae opposite which they lie, but fusion may occur which obscures their segmental character. The white visceral rami of all thoracic spinal nerves and I, I1 & 111lumbar nerves may terminate in the ganglion at the point where they enter or send fibres up or down the sympathetic trunk. The sympathetic preganglionic neurons have short axons and synapse in the chain ganglion or in a ganglion some disttance away from the vertebral column. The postganglionic fibre is usually long. Other preganglionic fibres pass without synapses through ganglia of the coeliac plexus, located in the abdominal region in front of the lumbar vertebrae. The prevertebral ganglia of the coeliac plexus include the coeliac superior mesenteric and inferior mesenderic ganglia. Preganglionic fibres pass directly to the medulla of the adrenal gland. This ectodermal, glandular structure, derived from neural-crest cells is composed of specialised, or modified, sympathetic ganglionic cells which are homologous with postganglionic sympathetic neurons and secrete adrenaline and noradrenaline too. Because adrenalineand noradrenaline serve as chemical signals of the postganglionic nerve endings, it could cause confusion in the adrenals which secrete them as hormones. Therefore, postganglionic fibres are absent. Since preganglionic nerve endings secrete acetylecholine, direct innervation of the adrenal removes the possibility of any ambiguity. Functions of sympathetic system Constriction of cutaneous blood vessels, causing pallor. Contraction of pili muscles, causing "goose flesh" and causing the hair to stand erect. Secretion of sweat glands. 4. Dilation of pupil. 5. Reduction in amount of saliva secreted. 6. Acceleration of heartbeat. 7. Dilation of the bronchi. 8. Relaxation or inhibition ofthe smooth muscles of the digestive tract. 9. Relaxation of bladder musculature. 10. Contraction of the sphincter muscles of the bladder. 1 1. Increase in blood sugar, red corpuscles in the blood stream. 12. Rise in blood pressure. 13. Decrease in cldtting time of blood. The above reactions together are usually associated with pain, anger, fear and gear up the body to react suitably to these situation. Parasympathetic nervous system The term 'craniosacral' outflow is frequently used to designate the complex of preganglionic fibres of the parasympathetic nervous system as the parasympathetic fibres depart from the v,vi, ix, and x cranial and spinal nerves from the sacral region.The trigeminal, oculomotor, facial, glossopharyngeal and vagus nerves are composed at least in part, of preganglionic parasympathetic fibres (Fig.10.8).The ganglia in which they terminate are situated close to or in, organs supplied by this system. Hence the preganglionic fibres are rather long and the postganglionic fibres are very short. The part of the parasympathetic system known as the sacral outflow is composed of efferent fibres which course through the white visceral rami of the 11,111& IV sacral nerves, which togetther form the pelvic nerve. The pelvic nerve supplies the lower part of the large ihtestine, kidneys bladder and reproductive organs. Postganglionic fibres within these organs are relatively short. Functions of the pamasympatheticsystem: 1) Dilation of blood vessels (except the coronary vessels of the heart). 2) Constriction of the pupil. 3) Increase in salivary and gastric secretion. 4) Constriction of bronchi.
- 25. ; 5) Contraction of walls of the digestivetracts. 6) Contraction of bladder musculature. 7) Relaxation of the sphincter muscles of the bladder. . 8) Dilation of blood vessels of the externalgenital organs. These reactions when taken together are associated with sensations of pleasure or comfort and conserveenergy. The general scheme of arrangement of autonomic system is similar in all tetrapodsexcept that it starts from the representation in a primitive form in lower vertebrates and increases in complexity as the evolutionary scale is ascended. Nervous Systemand Sense Organs SAQ 4 a) i) Which of the cranial nerves are purely sensory in nature? ii) Which part of the peripheral nervous system controls visceral activity? b) A nerve carrying information about the condition of internal viscera to the central nervous system is a .......................nerve. c) Which of the activitiesgiven below are controlled by sympathetic or parasympathetic part of the autonomic nervous system? i) dilation of pupil, iij increase in heartbeat iii) constriction of bronchi, iv) contraction of bladder muscles, v) increase in blood sugar, and vi) decreaseclotting time of blood. 10.5 BRAIN - A COMPARATIVE ACCOUNT You learnt in earlier sectionsthat all vertebrates share the major brain divisions, ten or more cranial nerves, spinal nerves and major spinal pathways to and from the brain. As we examine the chordates from amphioxusto mammals we find that the structureof brain undergoes variousmodificationsalongwith the evolution of major head sense organs(eyes, ears, nose, taste and lateral line system). The brain acquired major functions that were not found in primitive forms. As we comparethe brains of various vertebrateclasses (see Fig. 10.10)we find that the hindbrain has become specialised for processing sensations from touch, taste and balance from the near environment while the mid- and forebrain have become specialised for processing sensations from the eyes, and nose from the distant environment. 10.5.1 Jawless Fishes The brains of lamprey (Fig. 10.10a) and hagfish have a well developed hindbrain that suggests that its functionsare the most important. The cerebellum is small and the forebrain is mostly concerned with olfaction. This suggests limited 'locomotor abilities but highly developed sensory abilities, which is what is required in their environment. 10.5.2 Jawed Vertebrates The medulla is well developed in alljawed vertebrates showing its connectionswith the visceral network and as a screen through which all the informationenters or leaves the brain. The cerebellum is distinct in cartilaginousfishes (Fig. 10.10 b) with one or more transverse fissures. In teleosts the cerebellum is large in actively swimming fishes and relatively small in inactive fishes. Amphibians often have small or rudimentary cerebellum (Fig. 10.10d) reflecting simple locomotry abilities. In advanced tetrapods, the Iateral part ofthe cerebellum expands to control the muscles of the appendageswhich are specialised for locomotion. Cerebellum is seen in alligatorsamongst reptiles, and becomes more prominent in birds and mammals, again reflecting their complex locomotor abilities.
- 26. Functional Anatomy of Chordates - 11 Epiphysis .Tectum cerebellum cerebrum Medulla cerebrum T'ectum Medull* I Vagus Trctum Olfactotybulb , Medulla Vagus Cerebrum Medulla (c) (a) Cerebellum Cerebrum Teqtum Cerebellum Optic lobe - (0 Olfactory bulb Medulla Vagus. , (8) Fig. 10.10: Vertebrate brain (side view). a) Jawless fish. b) Cartilrginous fish. c)Bony fish. d) Amphibian. c) Reptile. f ) Bird. g) Primitive mammal. h) Advanced mammal. In mammals the sides of the cerebellum expand into separate hemispheres (Fig. 10.10 g). In primates, as in no other vertebrates the most lateral part is associated with finger coordination. The midbrain optictectum in particular is large in vertebrates that depend on their visual abilitiesbut in mammals it is relatively small as visual functions are taken over by the cerebral hemispheres. In primitive vertebrates the midbrain is relatively important as a principal centre of integration and its sensory linkto the cerebrum through thalamus is extremely well developed in amniotesespecially in mammals. The cerebrum is relatively small in all fishes. Olfactory bulbs and a thin walled pallium is present in cartilaginous and bony fishes. In fishes the cerebrum is mainly associated with olfaction. In amphibiansthe pallium is thicker than fishes. The firstchanga in the cerebral hemisphere is seen in reptiles with the migration of the inner nerve cells to the peripheral areas. 'The cerebral hemispheres enlarge greatly and grow backwards to coverpartially the diencephalon. The olfactory lobes merge with the cerebral hemispheres. In crocodiles the first emergenceof a true cerebral cortex or neopallium is seen.
- 27. In birds the olfactory lobes are 1,udimentary(Fig. 10.10 f ). Neopallium, however, is absent in birds. The cerebral hemispheres are large because the corpus striatum of birds is unusually large. In mammals, particularly in human beings the cerebral cortex is the most highly developed. The neopallium has taken over the greater part of the expanded and highly convoluted surface of the hemispheres and because of this expansion,the hemispheres tend to cover the other brain structures (Fig. 10.I0 h). Beginning with marsupials, a broad white mass the corpus callosum appears in mammals between the two hemispheres. It consists of medullated fibres and connectsthe neopallial cortical sides of the two sides. SAQ 5 Choosethe correct word from the parenthesis i) Olfactory lobes are well developed in (birds/mammals). ii) Neopallium is first seen in (birdslreptiles). iii) Cerebellum is most well developed in (reptiledmammals ). iv) Optictectum is not an important visual centre in (fishedmammals). 10.6 SENSE ORGANS In earlier sections you'learnt about the organisation of the nervous system of vertebrates. 'The peripheral nervous network gathers information from the various 'sensory receptors' of signals that reach the organism and these in turn are relayed to the central nervous system. In higher centres of the brain these signalsor impulses are interprstedas sensations.The receptor organs or 'Sense organs' themselvesdo not perceive any sensationsbut merely serve as means of accessto the nervous system.The information gathered by sense organs provides the body with a continuous preview of a changing environment. The information may concern stimulus quality (.eg. yellow light, static pressure, sweat,taste, pain, etc.), stimulus intensity (e.g. brightness, how strong) spatial patterns(orientation, distribution). Sensory receptors may be exo receptors or external receptorsthat receive informationfrom the external environmentor intero receptors that receive information from internalorgans. Broadly the various sensory receptors in the animal body can be classified accordingto the kind of energy they are able to perceive i.e. mechanical, chemical, light or thermal. For additional informationyou can refer to Table 10.2which lists the various kinds of receptorsfound in vertebrates. Table 10.2 :Extero and intero receptors of vertebrates. External senses Sight Photoreceptors Hearing Phonoreceptors Smell Olfactoreceptors Taste ~ustatorece~tors Touch an go receptors Pressure Temperature Heat Cold Pain Mecanoreceptors Thermoreceptors Thermoreceptors Caloreceptors Frigidoreceptors Currents of water ~l~esirece~tors Rheoreceptors Internal senses Muscle Position Propriortceptors Irr general most vertebrates have sensors for the five major sensesof taste touch, sight smell and hearing. Some vertebrates have greatly refined one or more of these familiar Nervous Syatem and Senae Organs
- 28. Functional Anatomy of ,! Chordates -I1 five. Let us first examinethe specific sense organs associated with sight, smell and hearing before we take up the special sensors. 10.6.1 The Eye The sensing of light is an important ability of chordate. Their most important photoreceptors are eyes, highly specialised structuresthat originate,as outpocketingof the brain. The simpilest and smallest vertebrate photo sensory organ is the median eye or parietal eye located near the middle of the top of the head (Fig. 10.11). Today only a fev fishes, and lizards possess a median eye which forms from the diencephalon.The lizard median eye consists of severalthousand sensory cells that transmit information to the brain. There is tran6parent lens overlyingthe sensory layer and light is concentrated by the lens on the sensory layer. The median eye is really a dosimeter of light exposure and does not produce any images as the lateral eyes do. Cornea . Lens Fig. 10.1 1 :a) Median eye in a reptile. b) sagittal section of median eye of reptile. We will discuss the human eye as an example of the vertebrate eye because eyes of all vertebrates are built on the same general pattern with variation accordingto their habitat. You would recall the development of the vertebrate eye from unit 17, Block -3 of the Development Biology course (LSE-06). The embryo~iic sourcesof the eye are the anterior brain or diencephalon for the retina and optic nerve ;ectoderm for lens and part of cornea; and nearby mesohrm for sclera,muscles and adjacent tissue. The first signs of the eye appear as lateral bulges of the diencephalo~i -the optic vesicles, with a small connection to the brain, the optic stalk. As the optic vesicle enlarges it contactsthe ectoderm of the head and invaginatesto form the two layered optic cup. The inner wall of the cup develops into the sensory retina, while the outer one forms the pigmented layer of the retina (choroid tapetum ) The opening of the cup narrows to form the pupil. The ectoderm thickens, invaginates to form the closed lens of the eye. The distal rim of the optic cup forms the ciliary body and iris along with the pupil. They are innervated with autonomic postggnglionicfibres. In addition to the ectodermal coat of the eye, a vascular choroid wall fused with the outer protective scleroticcoat is present. This entire structure is called the eye ball. In the front region the sclerotic layer becomes transparent forming the corneato whieh the conjuntiva is attached.The cornea has a complex development that includes scledal connective tissue, ectoderm, and neural crest cells. Cones are atlensttwo order of magnitude lens sensitive than rods to The eye functions like a camera and the retina is the screen on which the image is light and fail to function at nightor focused. This screen is multilayered and includes sensory and nonsensory cells. The in low intensity light. photosensitive cells are of two types, the rod cells and cone cells.Atthe margin of the retina near the ciliary body and iris there are no rods or cones.The rods contain a photo sensitivepigment rhodopsin or visual purple which gets bleached into lumi - rhodopsin by low intensity light and initiates rod cell activity to produce a visual 62 stimulus. In the cones another pigment iodopsin is present that is bleached only in high
- 29. intensity light. Vertebrates that usually live in low light levels or are nocturnal have more rods than cones. Those that need to see more details have large retinas because resolution is controlled by the density of receptors. Cone cells are responsible for colour vision. rSciera Suspensory ligament Anterior chamber Posterior chamher ciliary hody - d - _ _ - Extr~nsic eye muscle Chamberof the vitreous humor B!ind spot ,... a - Extrinsiceye muscle Fig. 10.12 :Structure of the human eye.The eyeball is allnost spherical in shape and is built of three layers I) The outer tough selerr that provides support and protection,2) The middle choroid coat containing blood vessels and nourishment,3)The light sensitiveretina. Several types of neurons in the retina convey information to the brain either directly or by interaction modify the input to the visual centres of the brain (see Fig.lO.13). Light energy is converted to electrical signals and transmitted by the bipolar cells to the ganglonic cells which then transmit the signal to the brain. Other neurons like amacrine and horizontal cells interact with the transmission between photo receptors and bipolar cells or between bipolar and ganglonic cells. The axons of the ganglion cells come together along the inner surface of the retina and turn inwards in one place to form a nonsensory area the blind spot and continue to the brain as optic nerve. Fibres from left and right optic nerves cross at the optic chiasma and the nerve impulses travel from right to left field of vision of occipital cortex. Light Hnrizontal cell Optic nerve Pigment epithelium Hnrizontal Light 0 Monosynaptic b~polar cell Fig. 10.13 :Photosensitivereceptors and other neurosensory cells of the retina. At the base of the retina is the pigmented choroid. It is either black or very shiny. Choroid pigment in day adapted animals is black and absorbs any stray light so that it does not get Nervous System and Sense Organs
- 30. Functional Anatomy of Chordates - Il reflected to the sensory retina. Reflected light produces images that do not get correctly aligned to the primary image and thus reduce visual acuity. Nocturnal animals have a mirror like choroid known as tapeturn lucidurn that reflects back the light into the retina. Because little light is lost through absorption by choroid their eyes are sensitive to dim light but at the cost of visual acuity because reflected image does not coincide exactly with primary image. Image formation in vertebrates is done by the cornea and crystalline lens. Cornea is almost flat in fishes but in terrestrial animals the cornea is curved and main image former and the crystalline lens is used for fine focusing. Many animals have a method to control the amount of light entering the eye. Like the aperture of the camera, their pupil which is an opening in the iris regulates the light intensity (Fig. 10.14). In most diurnal animal the pupil tends to be circular and relatively small. Nocturnal animals have round and relatively large pupils that permit maximum amount of light to enter the eye. Pupils of animals that are active both during day and night are able to expand greatly at night and become fine slits at day. A Dilated B ,C-at Shark tiom C Nocturnal Frog deep waters D Geko E Horse F House Cat C Human A noctbrnal Nocturnal Lizard carnivore Fig. 10.14 :'rhe pupils of most vertebrates are able to expand or contract to let more or less light enter the eye a) is dilated pupil in vertebrate to allow light, b-g) contracted pupils of vertebrates. As said earlier, the eye is protected by the tough sclerotic coat which merges with the cornea in front. Between the cornea and the lens is the fluid aqueous humor and between the lens and the retina is the fluid vitreous humor that help to hold the lens in place along with the muscle and are exchanged with blood on a controlled basis to supply nutrition to the lens. The sclera may contain cartilage or bone rings that prevent damage to the eye. The cornea is covered by a thin membrane known as conjunctiva. The eyelids and nictitating membrane found in many vertebrates help to protect and moisten the surface of the eye. Modified sebaceous glands secrete an oily substance that spreads on the cornea and lacrimal glands contain a watery fluid that lubricates, washes and moistens the conjunctiva. Comparative anatomy of vertebrate eye Fundamentally the vertebrate eye has the same plan. In some forms, the eyes are primitive, while in others they are degenerated and functionless. There occur variations in methods of accommodation, degree of retinat development and the pupil shape. The structural features of eyes among various groups are described below. 1) Cyclostomes In hagfishes the eyes lie beneath the skin, minute, degenerated and functionless. The cornea, sclera and the choroid are not differentiated. On the other hand, lamprey eye though primitive, is nevertheless a well-developed structure. The eyeball is flattened, sclera and the cornea are not fused to skin. There is a lack of suspensory ligament, and ciliary apparatus, the pupil is of fixed size and the eyelids are absent. Rods outnumber cones. 2) Fishes In elasmobranchs, the eyes, are large. In holocephalians the eyes are largest in relation to body size among all the groups of fishes. A characteristic optic pedicel is present. The sclera is cartilagi~ous and there is lack of intrinsic muscles in the ciliary body. In elasmobranchsthe surface of choroid coat contains light-reflecting crystals of guanin . Cones are absent in elasmobranchs. Colour v~sion seems to be wide spread in bony fishes,
- 31. Ciliary muscles and functional iris are absent. Deep-sea fishes have relatively enormous eyes. Recognition of enemies, prey, members of the same species and opposite sex is thus possible. In adult flatfishes, both eyes are on the same side of the head. The eyes of certain teleosts are adapted for vision in both air and water. 3) Amphibians In terrestrial amphibians movable eyelids, moistened with glandular secretions make their appearance for the first time. Closure of the eye is accomplished by retracting the entire eye within the orbit by means of retractor bulbi muscle. The protrusion of eye is brought about by means of levator bulbi muscle. The eyes in anurans are best developed amongst all the amphibians. No tapetum lucidum is present though the eyes appear to shine. It is doubtful if amphibians have colour vision.Ciliary bodies are'present. In caudate amphibians the eyes are small. The lids are absent in permanently aquatic forms. The caudate lens is exceptionally large and the ciliary body is less developed. The cave- dwelling salamanders show degenerated eyes. 4) Reptiles Eyes in reptiles show further adaptation to terrestrial life. Except for snakes and a few others, eyelids havebecome more movable. A true nictitating membrane and Harderian gland is present lacrimal glands are well developed except in snakes, chameleon and Sphenodon. A relative increase in number of cones is apparent. In most reptiles in the retina, a central area for acute vision is present. Colour vision is believed to exist in turtles and lizards but is of doubtful occurrence in crocodiles and snakes. In snakes, a fixed transparent skin is formed over the eyes as the eyelids are fused, A very important difference in the reptilian eye is the special ciliary apparatus, which alters the shape of the lens and corilea. 5) Birds The birds have uniocular vision except in birds of prey such as owls and to a lesser degree, hawks and eagles which have binocular vision. They have unifomi eye structure. The eyeball is very large and is correlated with aerial mode of life. The eyeball is partly concave al~d a highly developed nictitating membrane is present which offers protection during flight. The ciliary bodies are well developed. The cones predominate in birds of diurnal habit, while the rods predominate in nocturnal forms. Colour vision is wide spread. Pecten a special feature of birds is a serrated, fan shaped structure which extend into the vitreal cavity is well developed which might aid in perception of movement and may act as a supplemental nutritive device for retina. 6) Mammals 'The human eye structure is more or less typical of that seen in mammals. Variations are seen among mammals that are aquatic, terrestrial and those that lead an aerial life. Tapetum lucidum is present in nocturnal forms. Round pupil is most common but variations do occur. The retina mostly contains rods and cones. Though capacity of colour vision is mostly limited to higher primates. The optic nerve deccusation leads to binocular vision giving a third-dimensional effect. 10.6.2 The Ear The vertebrate ear is a specialised receptor for detecting sound waves in the environment. It usually functions in a dual capacity, serving at least in higher forms, both as an organ of hearing and of equilibration. What we see as the ear in mammals is actually only the external ear. The actual function of hearing and equilibrium are performed by the internal ear which has a similar structure in all vertebrates and is enclosed in a bony skull protected from the external environment. During evolution the vertebrate internal ear originated primarily as an organ for balance, the labyrinth also known as the vestibular apparatus. We will explain the structures responsible for equilibrium first. Vestibular apparatus In alljawed vertebrates the labyrinth has a similar structure. It consists of two chamber - like enlargements, an upper utriculus (little bottle) and a lower sacculus (little sac). These chambers are connected by a constriction the sacculoutricular duct. A narrow endolymphatic duct joins either the sacculus or sacculoutricular duct. Three narrow tubes, Nervo~~s System and Sense Organs
- 32. Functional Anatomy of Chordates - I I the semicircular ducbs, connect at both ends with utriculus (Fig. 10.15) and lie at right angles to each other. One of these lies in the horizontal plane while the other two are vertical, one directed forward and the other backwards. In cyclostomes the hagfish has only one semicircular canal which bears an ampulla at each end. In lamprey there are two semicircular canals. From fishes onwards all vertebrates have three semicircular canals and a slight projection of the ventral wall of the sacculus may be present. It is referred to as the lagena. The lagena is the forerunner of the auditory portion of the ear. Endolymphaticduct , > Macula utriculus Saccul~rs Lagena hlacula sacculus Co~nrnora canal AnteriOr semicircular Hagfish Lamprey (b) Fig. 10.15: a) Ceneralisrd vestibular apparatus o f vertebrates sl~owing the three sen~icircularcanals and major comprtments, the utriculus, sacculus andlagena. b) Inner ear of hagfish and lamprey. Wall of a~npulla Cupula Hair cell - Nerve fibers cells Otoconia Fig. 10.16: Sensory rbeptors i n the internal ear. a) One crista resides at the base o f each semicircular canal i n the ampulla. b) Macula containing otoconia reside in the three compartments of the inner ear. The membranous labyrinth is filled with endolymph a fluid that is more viscous than water. Almost completely surrounding the labyrinth is the perilymphatic space filled with a fluid perilymph, with is actually the cerebrospinal fluid. Surrounding the perilymphatic
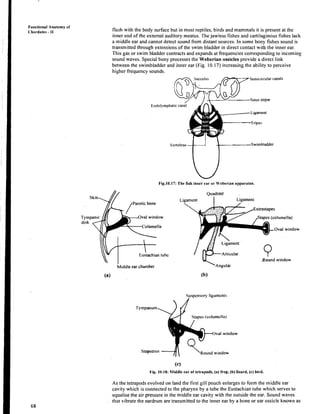
- 33. iI I ! space is cartilage or bone dependingon the species. In higher forms, a bony labyrinth, situated in the temporal bone, enclosesthe membranous labyrinth. The semicircular canalsare those portionsof the bony labyrinth, which surround thg semicircular ducts. The actual receptors for the sense of equilibrium consist of patches of sensory cell of cristae and maculae The former are located in the ampullaeof the semicircularducts and are made up of supporting cells and hair cells (see Box 10.1).The maculae lie in the walls of the utriculus, and the sacculus.The macula too is made up of a gelatinous cupula and hair cells except that embedded in its membrane is a high density mass of calcium carbonatecrystals known as otoconia (Fig.10.16). The semicircularcanals are designed to respond to rotational acceleration and are relatively insensitive to linear acceleration. When the head is rotated the fluid in the canal tends not to move at first because of inertia. Since the cupula is attached,the free end is moved in a directionoppositeto the movement of heads, stimulating the sensory hair cellsthus settingup impulses transmitted to the brain by branches of the auditory nerve. While rotational movements affect the cristae in the ampullae of the semicircularducts, the utriculus and sacculusare staticbalance organs that give information about the position of the head or body with respect to gravity. As the head is moved the stony mass moves over the hair cells sending nerve impulsesto the brain. Box 10.1: Detectingwater currents, maintaining balance and hearing sound seem to be very different sensory functions. However, all three are based on mechanoreceptors, sensory cellsthat respond to changes in pressure or mechanical force. The basic mechano receptor is the hair cell which has nothing to do with hair but is a cell which has a tiny hairlike process on its apical surface. Hair cells are transducers that change mechanical stimuli into electric signalsand these cells originatefrom surfaceectoderm. Each hair cell is embraced by a sensory fiber of neurons sensitiveto ionic changes in the hair cell. Through synapsesor similar contact points electrical excitation is passed on from hair cell to their embracing neuron which sendsthe signal on to the central nervous system. A neuromast organ is a small collection of hair cells, supportingcells and sensorynerve fibers and the projecting hair bundles are usually embedded in a gelatinous cap called cupula.The neuromast organ or its modification is the fundamental component of all three types of mechanoreceptive systems,the lateral line systems, vestibular apparatus that senses equilibrium and auditorysystem that responds to sound. -'4'. Hearing Hearing in vertebratesprobably appeared as a mechanism to alert animals to nearby activity that could be dangerous. Later it also became important in the search for food, mate and in communication.111 most vertebrates, part of the inner ear became modified to receive sound waves and certain hair cells in the inner ear became specialised to detect sound. You have learnt that the sacculus is fishes gives rise to a tiny pocket the lagena that, during evolution of vertebratesdeveloped into the hearing apparatusof the tetrapods. This lagena elongatesand in birds and mammals becomes coiled to form the cochlea. Within the lagena or the cochlea lies the sensory receptor for sound,the organ of Corti that is a specialised strip of neuromasts connected to the nervous system via the auditory nerve. The ear is made up of three compartments: external,middle and internal ear (Fig. 10.19 a) shows the typical mammalian ear, which is made up of all three compartments. lie external ear is absent in fishes and amphibians. It appears for the first time in reptiles in some lizards and crocodiles and is made up of a short tube the external auditory meatus that opens to the exterior by an externalorifice. In birds and mammals, the external auditory meatus is elongated.The part we consider as 'ear' is the pinna found only in the mammals. The pinna helps differentiatesounds from various directions and channels them into the external auditory meatus. Paired ears provide stereophonic hearing just as the paired eyes provide stereoscopic vision. The middle ear is made up of a diaphragm the tympanum or tympanic membrane and appears first in ancient amphibians. In amphibiansand a few reptiles the tympanum is Nervous System and. Sense Organs
- 34. Fuactional Anatomy 01 ' Chordates - I1 flush with the body surface but in most reptiles, birds and mammals it is present at the inner end of the external auditory meatus. Thejawless fishes and cartilaginous fishes lack a middle ear and cannot detect sound from distant sources. In some bony fishes sound is transmitted through extensions of the swim bladder in direct contact with the inner ear. This gas or swim blladder contracts and expands at frequencies corresponding to incoming sound waves. Speclialbony processes the Weberian ossicles provide a direct link between the swirnbladder and inner ear (Fig. 10.17) increasing the ability to perceive higher frequency sounds. Sacculus Semicircularcanals Sinus irnpar Fig.10.17: The fish inncr car or 'ehcrian apparat~rs. Middle ear chamber (a) Quadrate Ligament Ligament 1 . Extrastapes ,Stapes (columella) 1,igament Articular ~ A n g u , A r Round window Stapcs (columcllii) Oval window Round window (c) Fig. 10.10: hliddle car ol'tctrapods. (a) frog. (b) lizard. (c) bird. As the tetrapods evolved on land the first gill pouch enlarges to form the middle ear cavity which is connected to the pharynx by a tube the Eustachian tube which serves to equalise the air pressure in the middle ear cavity with the outside the ear. Sound waves that vibrate the eardrum are transmitted to the inner ear by a bone or ear ossicle known as
- 35. columella which first appears in amphibians (Fig. 10.18). Columella is a derivative of the hyomandibular of fishes. In some amphibians, reptiles and birds the collumella is tipped with a cartilaginous structure the extracolumella which rests on the undersurfaceo f the typlnpanic membrane. Nervous System and Sense Organs In mammals there are three bones in the middle ear: the stapes (stirrup) which is the reduced coluniella o f reptiles, the incus (anvil) and malleus (hammer) which are derived from the quadrateand articular bones respectively. These three bones form a chain that bridges the gap betweenthe tympanum and the inner ear (Fig. 10.19b).The middle ear is capable of transforming pressurewaves from the distance environment into mechanical motion. Semicircularcanals ~ditol.y nerve Roundwindow (a) Incus Malleus / / Tympanic membrane '1.0 auditory nerve .... .... .... ....:... 1 (b) Tectorial membrane - - Organ of Corti Fig.lO.19: Ear of mammal. a) external, middle and internal ear. b)The three middle ear ossicles. e) Internal ear. Note that the lagena has lengthened and coiled to form the cochlea You already know that the inner ear includes the vestibular apparatus and'surrounding pt.rilymphatic space. The auditory apparatus in the inner ear consists of the lagena in amphibians and reptiles. This lagena extends to form the tubular lagena in birds. In ~nammals the lagena forms the coiled cochlea. The organ of Corti is present along a central channel suspended within the lagena Two parallel perilymphatic channels run on either side of the central channel. The cochlea of mammals is coiled making two and a half turns in humans and is made up of three tubular canals running parallel with one another. These canals become progressively smaller from the baseto the tip (Fig. 10.19~). The three canals are, vestibular canal, the base o f which is closed by the oval window or fenestra ovalis. The end of the stapes expands into a plate whicli occupies the oval
- 36. Functional Anatomy of Chordates - I1 In neurophysiologya nucleus is a small cluster or aggregateof nerve cell bodieswithin the central nervous system. window. The other canal is the tympanic canal that is in communicationwith the vestibular canal at the tip of the cochlea and at its base is the round window or fenestra rotunda. A flexible membrane covers this window. Between these two canals is the middle canal or cochlear duct, which contains the actual sensory apparatus the organ of Corti. Within the organ of Corti are fine rows of hair cells that run lengthways from the base to the tip of cochlea. There are atleast24,000 of these hair cells in the human ear. -Eachcell is connecaedwith neurons of the auditory nerve. The hair cell rest on the basilar membrane which separatesthe tympanic canal from the cochlear duct .The organ of Corti vibrates with the basilar membrane in response to sound waves. In some vertebratesthe hair cells of the organ of Corti are embedded in a firm plate the tectorial membrane. When a sound wave strikes the ear, the energy is transmittedthrough the bones of the middle ear to the oval window, which oscillates back and forth driving the fluid of the vestibular and tympanic canals. The fluid oscillationsalso cause the basilar membrane with its hair cells to vibrate simultaneouslycausing impulses to be generated that are carried by the auditory nerve to the brain where they are interpreted by the hearing centres. The loudnttss depends on the number of hair cells stimulated and the quality of tone is produced by the pattern of hair cells stimulated. 10.5.3 Olfactory Organs The sense of smell or olfaction involves chemoreceptorsusually located in the nasal passages. There are three components involved in olfaction. The olfactory epithelium, which is specialised epithelium within the nasal cavity. It contains basal cells, supporting cells known as sustentacular cells and the olfactory sensory cells which are the actual chemoreceptors(Fig. 10.20).Each sensory cell has a tuft of cilia at its apical end, embedded in mucous secreted by the sustentacularcells and they communicate directly with the olfactory bulb by their axons that pass through the bony cribriform plates. Fig. 10.20 :Cellular organisationof olfactory epithelium. The sensory cells are actual nerve cells that penetrate the epithelium with modified dendrites. In most fishes the olfactory sensory receptors are embedded in paired blind-ended pits known as nasal sacs. Water carrying chemical flows in and out of these sacs as the fish swims. In tetrapods, a small external opening or naris provides access to each nasal passage and the black of the nasal passage opens into the mouth through the internal naris. In amphibians,some reptiles and mammals a separate and distinct region of the olfactory epithelium formsthe vomeronasal epithelium of the vomeronasal organ or Jacobson's orgah (See Fig. 10.21). The vomeronasal organ is absent in most turtles, crocodiles, birds, some bats, primates and aquatic mammals. Vomeronasal organ is an accessory olfactory system and its sensory cells project into the lumen by means of microvilli. The neural circuiting also is separate and runs parallel to the olfactory neural system. The sense of smell is well developed in fishes but is a secondary sense in bats, birds and primates. One way flow of water in cquatic vertebratesensures a continuous flow of new water to wash away the chemicalsthat have been detected by the olfactory epithelium. In tetrapods air replaces water and air that flows through the nostrils must pass the olfactory epithelium on its way to the lungs. The sniffingof terrestrial vertebrates increasesthe turnover of air in the nasal chamber and thus the animal can sample the environmental odors.
- 37. Nervous System and Sense Organs Nasal sac 2 Nonchoanatefish 1Nasalsac I I Extmal naris Nasal chamber ! I iternal naris Internal naris lnternal naris Choanate fish Mammal Nasal sac External naris Internal naris Amphibian Fig. 10.21: Olfactory organs andvomeronasal organ in vertebrates. Note that the vomeronasrl organ is absent in fishes. SAQ 6 a) What is the function of rods and cones? b) The space between the membraneousand bony labyrinth is filled up by .................. C) Which part of the ear is responsible for the sense of balance. d) Name the three bones in the middle ear in mammals. e) Lagena is present in mammals in the form of ...................... f) What is the function of the parietal eye? g) What are the sensory cells in the ear responsiblefor hearingcalled? h) What are the gravity sensors in the inner ear called? In the next section we will discuss some specialisedsense organs found only in particular groups of vertebrates. 10.7 SPECIALISED SENSORY ORGANS Some sensory structures are considered to be so unique, or so complex that they are called special sensors. for example electroreceptors, lateral line system, pit organ thermoreceptersof snakes and system for ecolocation. We will first discussthe lateral line system in fishes. 10.7.1 Lateral Line System in Fishes Reception of deep vibrations in water and of stimuli caused by currents or movements of water, including minor currents produced by the animal itself, are amongthe functions ascribed to the lateral-lineorgans. The lateral line system is present within the skins of most cyclostomes, other fishes and aquatic amphibian,but is unknown in terrestrial
- 38. Functional Anatomy of Chordates - I1 vertebrates The sensory receptors of the lateral line system are the neuromasts(referto Box 10.1). The role of neuromasts of fishes has remained controversial. However, it is supposed to have a secondaryauditory function,temperature reception and a chemical sense. Disturbances of mapy kinds in the surroundingsof a fish can activate the neuromasts, but water moving on the body surface provides most active stimulation. The sensory hair can detect two types of water currents i.e. water moving from head to tail and from tail to head. Weuromastssend a steady spontaneoustrain of bioelectric potentials along their nerves and the rhythm of the pulse changes during stimulation. The sensory cells are pear-shaped and have hair-like processes at their free ends (Fig. 10.22c). The neuromasts are supplied with Vllth and Xth cranial nerves. The canals contacting the neuromasts are in the form of open grooves in primitive shark, while in Holocephali,they are fully exposed. Majority of elas~nobranchs and teleosts contain closed tubes filled with mucous. 'They open to the exterior at intervals by means of tubules (Fig. 10.22 a). Lateral line system is sensitivet o surfacewaves and concentric ripples, allowing fishesto detect prey in the water. Removing the lateral line organs or blocking them causes disorientation in fishes and loss of ability for schooling behaviour. , Ncrvc Ki~lociliuln Stereocilia Skin surface Hair cell Support cell Sensory llervc Motor nerve Fig. 10.22: a) Lateral line organs in fish, b) canal organ neuromast. Each opens to the exterior by a pori. c) superficial neuromast. Electroreception in Fishes Besides functioningas an organ for detection of waterturrents, there is a considerable agreementthat a part of neuromast system responds to weak electric potential. This information helps the fish to orient itself, to avoid enemies and to participate in schooling. Most fishes produce enough electric potential when their muscles contract to make their presence detectable,Others produce a more powerful electric field by specially modified muscles or tissues-the electric organs and can monitor disturbances in their field caused by objects that enter it. Fishes that have electric organs may use them to signal other individuals or to defend themselves. Electroreceptorsare not effective in terrestrial animals, as electriccurrents are not conducted so easily in air as in water. 10.7.2 Pit Organs in Snakes 'The pit receptors of'reptiles bear a striking morphological resemblanceto the external neuromasts of fishes. Hence it has been hypothesised that the pit organs are evolutionary
- 39. derivatives of neuromasts. However, their innervation by spinal or Vth cranial nerve is not consistent with this hypothesis. Two families of snakes, Viperidae and Boidae have specialised heat sensorsor receptors on the body surface in the form of pits. Pits open to the surface between epidermal scales. These pits contain free nerve endings that are excited by infrared radiation and the information is transmitted to the optic tectum of the midbrain. Apical pits are scatteredon the body surface, mostly on the trunk, and provide sites for the input of tactile stimuli. In boa constrictors and pylhons of family Boidae the free nerve endings lie between epidermal scales along the lips these are known as labial pits. In sub-familyCrotalinaeto which the snake, pit viper belongs, specialised pit receptor is found on the head. The pits are located on the posterior region of loreal scale, which is located between the external nares and the eye, and are termed as loreal pits or facial pits. These pits are directed forward and may be several millimetres wide and equally deep (Fig.10.23 a and b, see also Fig. 3.31, Unit 3, Block 1). These differ from the labial pits of pythons. Sensory nerve endings are suspended.in a thin pit membrane halfway between the bottom of the pit and not lying at the bottom as in pythons. The pit or cavity is divided into an inner and outer chamber.A duct between the inner chamber and the skin of the snake may prevent differentialchanges in pressure arising between the two chambers. The separating membrane is innervated by trigeminal nerve, which is responsible for the input from the head sensorto brain in snakes. Experimentally it was recorded that the frequency of nerve impulses,increasesas the receptor object warms up. Fluctuation occurs during the cooling of the object that is registered by the pit. Fig. 10.23: I'hermoreceptive pit organs In snakes. a) Facial pits in viper, b) Labial pits in python. Since birds and mammals are warm bloodied, they emit infrared radiation and experiments show that temperature changes of 0.003°C can be detected by the pit organs. It was observed that a consistent change in the pattern of electroencephalogramoccurs after a stimulus has been received by the peripheral nervous system called as evoked potential. It was also shown that the mitochondriaenlarge after exposure of the receptor to an infrared stimulus, while when exposed to cold body, the mitochondria are condensed. 10.7.3 Echolocation in Bats Nervous System and Sense Organs ~mong'st mammals bats and cetaceans have made hearing as a distancesense for navigation. It was found that insectivorousbats produce sound waves of high frequency. The human ear cannot perceive these sounds. The bats perceive objects by emitting sound waves that are reflected by the objects and are then detected by the bat's ears. Bats can thus locate and avoid obstacles in total darkness by means of echolocation.
- 40. FunctionalAmatonly of Chordates-ll The vespertilionid bats produce very short sound impulses each lasting only approximatelyone to two milliseconds. These ultrasonic bursts are produced in the larynx and emitted through the slightlyopened mouth. The animals detect the difference between the emission of a sound and its return as an echo. The time differential in the arrival of the echo into the differentears gives information about the direction of the reflectingobject. The horseshoe bats (Rhinolophidaeand Hipposideridae)have peculiar cutaneous growths on their noses (Fig.10.24) and emit ultrasonic sounds of consistentlyhigh frequency. They can produce sound with the mouth closed or while catching and eating insects. The horseshoe-shaped flaps around the nostrils are used as a sound cone, almost as a megaphone, during the emission of ultrasonicsounds; and the curvature of these flaps can be changed by musale contraction.Thus, the animalsare in a position to change the width of the sound cone in accordancewith the different distances of the sound reflecting object by sweepingthe sound beam back and forth to scan their surroundings . The cave dwellingmembers of the genus Rousettus orient themselves with the help of large eyes when sufficient light is aiailable. In total darkness,they can produce ultrasonic t1 cries and use them for, echo sounds. 'These sounds are not made in the larynx, but by a clicking of the tongue. The most surprisingof all the specialisedbats are the speciesthat feed on fish. These bats have a well-developed system of frequency-modulatedsonar, but sound loses much of its t energy in passing frtim air into water and from water to air. ~ c h o Pulse Fig. 10.24 :a) Echolocation of an insect by little brown bat. Myotis lucifugus. b) Modification of nose to broad cast the FM pulses. SAQ 7 Fill in the blank spaceswith appropriatewords: ................. i) Lateral line organs consist of sensory papillae called which are composed of..,...............surrounded by supportingcells. ii) Pit organs of snakes are ...................receptors.
- 41. ... 111) ...................is a method used by bats to orient themselves in space by detecting the differences between the ..................of the sound and its return as an ............................ ..................... iv) The cave dwelling bat produces ultrasonic cries by 10.8 SUMMARY - - - ---- - After studying this unit you have learnt that: Almost all the nervous system is derived from the neural tube and neural crests which appear very early in embryonic development. Nerve fibres are of two types :medullated or nonmedullated. Medullated fibres are white in appearance; nonmedullated fibres are grey. Bundles of nerve fibres are called nerves, whereas aggregations of nerve cell bodies are known as ganglia. The nervous system is divided into three m a i ~ parts a) the central nervous system composed of brain and spinal cord; b) the peripheral nervous system, made up of cranial and spinal nerves and c) the autonomic system, consisting of portions of certain cranial and spinal nerves, as well as outlying ganglia connected with those structures of the body under involuntary control. In lower forms, brain and spinal cord are disposed in a straight line. In higher forms, flexures or bends occur which make possible the accommodation of a larger mass of nervous tissue with~n relatively narrow confines of the cranium. Nervous System and Sense Organs ( The greatest changes occur in the development of the telencephalon which is a part I of the forebrain. The anterior portion of the forebrain is primarily concerned with the olfactory sense. The floor of the diencephalon, called the hypothalamus, contains centres which integrate the activities of the autonomic nervous system with those of other nervous tissues. The anterior part of the roof of the diencephalon remains epithelial but forms the anterior choroid plexus, while posterior portion may give rise to parietal and pineal outgrowths. An invagination from the floor, the infundibulum forms the posterior lobe of the pituitary gland. The dorsal part of mesencephalon forms paired optic lobes. The anterior part of the haidbrain called the metencephalon, gives rise to the cerebellum which coordinates the neuromuscular mechahism of the body. The myelencephalon or medulla oblongata is the posterior p&t of the hindbrain. The roof remains epithelial and forms a posterior choroid plexus. Brain and spinal cord are surrounded and protected by membranes called meninges. The spinal nerves are paired. Most spinal nerves give off three branches of rami. ' The cranial nerves, arising from the brain, number 10 in anarnniotes and 12 in amniotes. Some cranial nerves are purely sensory (I, I1 and VIII, and other are purely motor (111, IV, VI, XI, XII). The remainder (V, VII, IX, X) are mixed (sensory and motor). The autonomic nervous system is made up of sympathetic and parasympathetic components. Each consists of preganglionic and postganglionic system. Preganglionic sympathetic system is often called the thoracolumbar outflow, while the parasympathetic system is referred to as the craniosacral outflow. Receptor organs are structures capable of responding to definite stimuli by setting up impulses which are in turn transmitted by nerve fibres to the centre nervous system. Receptor organs are classified as external and internal sense organs. The vertebrate eye has no true homologue among other phyla and appears for the first time in cyclostomes. In the vertebrate eye, light must pass through various cellular layers of the retina before it can stimulate the sensory receptors, or rod and cone cells. Six broad, strap- shaped muscles move the eye. Other accessory structures associated with the eyes of different vertebrates in each case serve to adapt the animal better to its own particular environmental needs. The vertebrate ear, at least in higher forms, functions in a dua! capacity as an organ of equilibration and hearing. The sensory portion of the ear is called the inner ear or membranous labyrinth. The portion concerned with equilibrium consists of utriculus, sacculus, endolymphatic du'ct and three semicircular canals. The portion concerned with hearing is lagena. In mammals and birds it is the coiled choclea.
- 42. Functional Anatomy of Chordates - I1 0. The receptors for the sense of smell are confined to the olfactory epitllelium of fishes, and olfactory epithelium of the nasal canal of tetrapods. In higher forms outgrowths from the walls of the nasal passages in the form of folds and scrolls (tubinates or conchae) increase the surface of the respiratory and olfactory regions. Neuromast organs are fluid-filled pits consisting of sensory and supporting cells and sensory terminals. They may open to exterior or occur located enclosed in canals. They are present only in fish and aquatic amphibians. Electroreception and mechanoreception are the attributed functions. Neuromast organs are innervated by cranial nerves V11, 1X and X. Snakes exhibit pit receptors of uncertain homology. Apical pits occur at the apex of body scales. Pit organs occur on the head of vipers and boas. Apical pits are probably mechanoreceptors, while pit organs are thermoreceptors. Echolocation is observed in bats. They make use of reflected sound waves in orientation. This method highly developed in rnicrochiropteran bats that emit high frequency and short wavelength sounds. 10.9 TERMINAL QUESTIONS 1. What are the primary divisions of the nervous system? Add a note on their subdivisions. ...................................................................................................... I 2. Name the flexures and their position, that are present in vertebrate central nervous 1: system. 3. Mention the cranial nerves of special senses and the nerves that innervate the eye muscles. ....................................................................................................... 4. What are th'ecomponents of the dorsal and ventral column in the spinal cord? ....................................................................................................... 5. Which part of the brain is well developed in all vertebrates and why? .......................................................................................................
- 43. 6. With which parts ofthe brain are the essential senses of small sight and hearing associated? ...................................................................................................... ...................................................................................................... Nervous System and Sense Organs 7. What are neuromasts? Give three functions of neuromasts. 2...................................................................................................... 8. What are pit organs in reptiles? I-low do vipers and boas locate the prey? 9. What do you understand about the pheno~ncnon of echolocation? ....................................................................................................... 10. What is electro-reception in fishes? Co~nment on its functional significance? 10.10 ANSWERS Self-assessmentQuestions 1. a) Brain is formed mostly of grey matter except the medullated tracts while spinal cord is formed of both grey and white matter. b) Myelin sheaths are secreted by Schwann cells and oligodendrites. c) Myelin sheaths are found both in brain and spinal cord. d) White and grey matter intermingled in the brain forrns the reticular formation. 2. a) Telencephalon matches lateral ventricles Diencephalon matches third ventricle Rhombencephalon matches fourth ventricle Medulla oblongata matches myelocoel b) i) sensory ii) dorsal roots iii) motor neurons iv) dorsal funiculi v) ventral funiculi 3. a) Metencephalon - cerebellum mesencephalon - optic tectum diencephalon - epithalamus, hypothalamus Telencephalon - thalamus, cerebrum rnyelencephalon - medulla oblongata
- 44. Functional Anatomy of Chordates - 11 b) i) menix primitiva ii) dura mater and pia arachnoid iii) dura mater, pia mater, arachnoid 4. a) i) 0,(Terminal); I, (Olfactory); I1 (optic); VIII (auditory) b) autonomic motor c) Dilation of pupils - sympathetic increase in heartbeat -sympathetic constriction of bronchi - parasympathetic contraction of bladder muscles -parasympathetic increase in blood sugar - sympathetic decrease in clotting time - sympathetic 5. i) mammals ii) reptiles iii) mammals iv) mammals 6. a) Rods are egceptionally sensitive to light receptors while cones are also photoreceptors less sensitive than rods. Cones are responsible for visual acuity and colour vision. b) perilymph c) inner ear or the vestibular apparatus d) Incus, stapes, malleus e) Cochlea f) It is a dosimeter for light and does not form images. g) Organ of Corti. 11) Sacculus and utriculus. 7. i) Neuromasts, hair cells ii) Infrared rays iii) Echolocation, emission, echo iv) clicking of tongue. Terminal Questions 1. The primary subdivisions of the nervous system arc: i) central nervous system and the ii) peripheral nervous system. The subdivisions of the central nervous system are composed of brain and the spinal cord and the peripheral nervous system consists of spinal nerves, cranial nerves and autonomic nerves. 2. Three types of flexures are present in vertebrates: i) a cephalic flexure between forebrain and midbrain, ii) a cervical flexure near thejunction of medulla oblongata and spinal cord. iii) A pontlne flexure in the region of metencephalon. 3. Terminal nerve, olfactory nerve and the optic nerves are termed as the nerves of special sense. Occulomotor, trochlear and abducense nerves innervate the eye muscles. 4. In the spinal cord, the dorsal column is composed of upper somatic sensory and lower visceral sensory regions. The ventral column in turn, is composed of upper visceral motor and lower somatic motor regions. 5. Myekencemalon or medulla is the oldest part of the brain and well developed in all vertebrates as it controls the vital functions of the body, including the involuntary functions and equilibrium. 6. Smell is connwted to the forebrain; sight to midbrain and hearing is connected to hindbrain. 7. Neuromasts are the sensory papillae surrounded by supporting cells, present on the surface of the skin. The three functions of neuromasts are: (1) short distance auditory function, (2) temperature reception and (3) chemical sense. 8. Pit organs in reptiles are the thermoreceptors that respond to radiant heat. Vipers and Boas can detect even very mild fluctuation in temperature and warm-blooded animals at a distance of several feet, and thus locate the prey. 9. Echolocation is the pbnomenon in which the animal can orient itself, avoid obstacles and also detect the prey with the help of reflected sound. 10. Electroreception is a special sensory system in fishes, through which the animal produces electrical potential and is able to detect, and paralyse the prey. It also helps them in schooling and breeding behaviour.
- 45. UNIT 9 URINOGENITAL SYSTEM Structure 9.1 Introduction Objectives 9.2 The Urinary System in Chordates Urinary System in Protochordates Urinary System in Vertebrates 9.3 Embryonic Development of Urinary System 9.4 Uriniferous Tubules 9.5 The Kidney Structure of Kidney Blood Circulation in Kidney 9.6 Phylogeny and Succession of Kidneys 9.7 Functions of Urinary System 9.8 Variations in the Urinary System Plan Habitat Related Structural Variations Variations in Urinary Systems of Vertebrates belonging to different vertebrate Groups 9.9 The Genital System Embryonic Origin of Gonads and Gametes Functions of the Genital System Genital System of Protochordates 9.10 Male Genital System'of Vertebrates Testes Male Genital Duct Male Accessory Sex Glailds IntromittentOrgan5 9.11 Female Genital System of Vertebrates Ovary Female Genital Ducts Female Accessory Glands External Female Genitalia Mammary Apparatus 9.12 Survey of Gonads in Vertebrates 9.13 Summary 9.14 Terminal Questions 9.15 Answers 9.1 INTRODUCTION - - - Excretion and osmoregulation as you are aware (Refer LSE-05 Unit 4) are two homeostatic processes performed by the kidneys and urinary ducts, which form the urinary system. This system is intimately associated both anatomically, and in terms of embryonic origin with the genital system. The genital system includes the gonads wlilcli generate gametes and the genital ducts that serve as passages for the gametes - sperms In males and ova in females. Though functionally different the two organ systems the urinary and the genital system are treated together as the urino- genital system, since both develop from the same segmental blocks of trunk mesoderm or ad-jacenttissues and share many of the ducts. Thus although the two systems have nothing common functionally they are closely associated in their use of common ducts and are studied under the broad heading of urinogenital system. In this unit, you will learn about the embryonic development of the urinogenital system, the basic plan in vertebrates and the variations in different chordate groups. You will also study the adaptive variations of urinary system in vertebrate living in specialised habitats. Objectives After going through this unit, you should be able to: describe and illustrate the urinogenital system of the proto chordates - cephalochordates and urochordates, illustrate and describe the basic plan of urinogenital system in vertebrates, give a comparative account of the urinogenital system of fish, amphibians, reptiles, birds and mammals.
- 46. Functional Anatomy of Chordates -11 Nucleus describe the origin and embryonic development of kidneys, gonads and their associated ducts, discuss the morphological and physiological adaptations of the urinogenital system of vertebrates. 9.2 THE URINARY SYSTEM IN CHORDATES 9.2.1 Urinary System in Protochordates The urinogenital system of protochordates such as the Cephalochordate [e.g. Branchiostoma (amphioxus)] ahd Urochordate, (e.g. Herdmania) is very different both in structure and origin from that of the vertebrates (Refer unit 1). Cephalochordata Despite the similarities to vertebrates in other aspects of its anatomy, the specialised organs of excretion in the cephalochordate, Branchiostoma (amphioxus) show no relationship to any part otthe vertebrate kidney or other known fluid regulating structure. The organs of excretion in Branchiostoma are the protonephridia -which are ectodermal in origin (unlike vertebrate kidneys which originate from the mesoderm). Protonephridia(Fig, 9.1) are segmentally arranged sac-like tubes which lie in coelomic spaces or atrium abbve the pharynx. Each protonephridium opens into the peribranchial or atrial space surrdunding the gills so that its excretory products released into the atrium, can be flushed away by the outgoing stream of water. Internally, within the coelomic sac the protonephridium terminates blindly (not opening) into the coelom. (see also Fig. 9.17a in this unit) Dorsopharyngeal coelom ./ Coelomic fluid II Fig. 9:l: The protonephridium of Broncl~iostoma. (Arrows show path of fluid flow) i The protonephridibl sac bears numerous, specialised tubular flame cells called the solenocytes (Fig. 9.2.). Each solenocyte has a single flagellum projectrng downwards into the protonephridial tube or canal. Constant beating of flagella probably forces fluid into the protonephridia. The exact mechanism of functioning of solenocytes is still not -Flagellum clear. As the distql ends of solenocytes lie close to branchial (gills) blood vessels, the solenocytes probably help to filter blood fluids from the branches of branchial blood vessels. In the prdcess some fluid components such as ions are probably returned to the body (Fig. 9.1). Fig. 9.2: Enlarged view of a single flame cell or solenocyte. Urochordata In the urochordatg, Herdmania, neural gland is supposed to be the excretory organ (Fig. 9.3) and lies embedded in the mantle above the nerve ganglion or brain. The neural gland consists of few central tubules from which arise peripheral tubules. The central tubules open into a longitbdinal canal. The longitudinal canal opens by a ciliated funnel at the base of dorsal tubercle. The excretory cells are called nephrocytes. They are present in blood in order to kollect waste which are in the form of xanthene and urates. The waste passes through th lumen of the neural gland and its duct and is finally discharged into the pharynx. Neufa1 glands are endocrine in function as well, since they also control oviposition, development and metamorphosis.
- 47. Zlrinogenital System Bloodsinusci hr,of ncunl u Fig. 9.3: Neural glar~d of Herdnranin. 9.2.2 Urinary System in Vertebrates The basic plan of urinogenital system is common to all vertebrates. The urinary organ- system lies in the abdominal region (Fig. 9.4a and b) and consists of paired kidneys which are the principal organs of excretion and osmoregulation. These kidneys lie dorsal to the coelom, (retroperitoneal), one on each side of the dorsal aorta. The basic units of the kidney are rhe minute, uriniferous tubules that end blindly and receive filterate from the blood. The uriniferous tubule consists of two parts, the nephron and collecting tubule. The primary function of the uriniferous tubules is removing excess water, salts, waste ' metabolites and foreign substances from the blood. Numerous such tubules connect to form an urinary or excretory duct system called ureter which carries urine from the kidney. The ureter of each kidney opens into a urinary bladder which in most adult tetrapods is a single median structure. Urinary bladder acts as a reservoir which stores urine prior to its removal. The bladder develops as a ventral outgrowth from the cloaca. The bladder opens to the body surface through a short tube, the urethra. The urethra is thus a small tube which arises from the bladder and opens by a single,urinary opening either into the cloaca or directly to the outside. l-------'II~l~qt~Ir b n w r ard Fig. 9.4: Ventral view ofthe human excretory systems in (a) male. (b) female. 9.3 EMBRYONIC DEVELOPMENT OF URINARY SYSTEM Urinogenital system is mesodermal in origin. The intermediate mesoderm which is located in the dorsal and posterior body wall of the embryo forms the kidney. At the onset of its differentiation this posterior region of the intermediate mesoderm expands to form a nephric ridge that protrudes slightly from the dorsal wall of the body cavity (Fig. 9.5 a). Next the paired nephrotome which are the embryonic fore runner of the nephric tubule (Fig. 9.5 b) are formed from the posterior part of intermediate mesodehn. The nephroto~ne is often segmented and soon differentiates into a series of segmentally arranged tubules each of which contain a nephrocoel, which is a coelomic chamber that
- 48. Functional Anatomy of Chordates- I1 opens into the coelom by means of a ciliated peritoneal funnel. The medial end of the nephrotome then widens to form a thin walled renal capsule into which enters a tuft of arterial capillariesaalled glomerulus. From the lateral end of the nephrotome arise outgrowths. 'These outgrowth fuse to form a common nephric dpct (Fig. 9.5 c). 'The nephrotome is now properly called a nephric tubule and may retain its connectionwith the coelom through the persistent peritoneal funnel. Thus the fundamental plan underlying the excretory system consistsof paired and segmented nephric tubules that open on one end into the coelom and on the other end into the nephric duct with a .glomerulus in behveen (Fig. 9.5 d). In most adult vertebrates,however, there is no connection with the coelom through peritoneal funnel. Mewneph~ duct In the development of the urinary duct of ureter, longitudinal ducts appear first at the anterior end of nephrogenous mesoderm as backwardly directed extensionsof the excretory or nephric tubules. The extensionsjoin to form the 'nephric duct' which opens into the cloaca. Al~l the nephric tubules ultimately open into this duct. tubule The numberofuriniferoustubules vary from only afew hundred in the kidneysof cyclostomes to over a million per kidney inmammals, in whom the tubules of both kidneys combined constitute over 120km of tubing. Nave l u k n ric Fig. 9.5: Embryonia developmentof nephric tubules in vertebrates. (a) Embryo (lower part) showing location of developing kidney (nephric ridge); (b) sectionthrough embryo showingthe appearance of segmental nephrotome in the posterior part of the intermediate mesoderm; (c) diagrammlatic representationof developing kidney tubule and longitudinal nephric duct. note the segmeatalarrangements; (d) the medialend of the nephrotomesdifferentiates into the first part oftha nephric hrbule. the renal capsule into which the glomeruls grows. Arterial sprouts from dorsbl aorta form the glomerules. The lateral ends of nephrotomesgrow outward and fuse with each ~ t h e r to form the nephric duct. Sometimes the nephrotomes remain connected to the coelom byi means of ciliated peritoneal funnel. The functional units of the kidney as mentioned before are the minute uriniferous tubules (Fig. 9.6). Each tubule is tonstituted of (i) nephron (nephric tubule which is the excretory unit) and the (ii) collecting tubule (Fig. 9.6) into which the nephron empties. Nephrbn (nephros: Gk Kidney); concentratesurine and passes it through the collectingtubule. The collecting tubule affectsthe concentration of urine and conveys it to the minor calyx, (see Fig. 9.9 a) which is the beginning of the excretoryduct.
- 49. Fig. 9.6: llriniferous tubule -the basic excretory unit. llrinogenital System I) Nephron Each nephron has: a) a dilated portion -renal (Malpighian) corpuscleand b) the secretorytubule portion. (a) Renal Corpuscle or Malpighian Corpuscle The renal corpuscle is the proximal portion of each nephron and is made of (i) double walled squamousepithelial capsulecalled Bowman's or renal capsule. The outer layer of the capsule is called parietal layer and inner one is called the visceral layer. Between the layers of Bowman's capsule is a urinary space receiving fluid filtered through capillary wall and visceral layer. (ii) A tuft of inter-arterial capillaries called glomerulus is surrounded by the Bowman's capsule. Each glomerulus is formed by the subsequentbranching of the renal artery (one of the major branches from the dorsal aorta) into a capillarity bed. The visceral or inner layer of the Bowman's capsule is made of stellatecells called podocytes. The visceral layer is in contact with blood vessels of glomerulus. The point of eMry of the blood vessels forms the vascular pole and the entry point of the nephric filtrate from the Bowman's capsule into the proximal convoluted tubule is called the urinary pole. Fig. 9.7 shows a complete uriniferoustubules,surrounded by its blood vascular system. (b) Tubular portion The tubule portion is subdivided into (a) proximal convoluted tubule (PCT) (b) loop of Henle with thick and thin limbs and (c) distal convoluted tubule (DCT).The PCT begins at the urinary pole of renal corpuscle(Fig. 9.8) and is Kited by simple epithelial cells which have microvilli forming a brush border and lots of mitochondria and the enzymeNa' K' ATpase . PCT has a wide lumen and is surrounded by peritubular capillaries. Henle's loop is U-shaped with thick descendingand ascending limbs. Initiallythick (the thick part is the straight part or pars recta of the proximal tubule)the loop of flenle thins near the turns then thickens again as it becomes part of the distal convoluted tubule. Henle's loop has a large lumen lined by squamous epithelium but no brush border. The distal convoluted tubule (DCT) is continued from the ascending limb of Henle's loop and forms the last segment of the nephron. It is lined by cuboidal epithelium which lacks brush border. At the vascular pole, DCT establishes contact with renal corpuscleof the parent nephron (see Fig. 9.7 a, b and 9.8).
- 50. Functional Anatomy of Chordates- 11 .t v v n t l layer made orpodtxytca I Rovrmm s apeprulc I Fig. 9.7: The basic excdetoryunit :a)uriniferous tubule showing its relationship to its surrounding blood vascular systelm, b) an enlargement of the boxedarea in a showing the juxta glomerular apparatus. 11) The CollectingTubule and CollectingDuct Urine passes fdom the distal convulated tubule to the collecting tubule. Collecting tubules of nephronsjoin together to form collecting ducts or papillary ducts of Bellini. These ductswiden neartips of renal or medullary pyramids of the kidney. Distal tubule Fig.9.8: Magnified view of the Borvman's capsule showing poles the jurta'glomerular cells and ~nacula densa.
- 51. Juxtaglomerular Apparatus llrinogcnital System 'The wall of afferent arteriole near the renal corpuscle is made up of modified smooth muscle cells called juxtaglomerular cells which contain lots of Golgi bodies. The secretion of these cells is Renin which helps to regulate blood volume. The wall of the distal tubule at this region is modified and looks dark under the microscope,hence it is called macula densa. The afferent arteriolejuxtaglomerular sells and macula densa form thejuxtaglomerular apparatus (Fig. 9.7 b). 9.5 THE KIDNEY - - The kidneys are specialised for maintaininghe appropriate levels of water and many solutes in the body and for eliminating wastesof protein metabolism and in canying out osmoregulation. The kidneys of vertebrates are especially important in eliminating excess water and divalentions and conservingsolutes. In this sectionwe willjust deal with the kidney and the urinary system of vertebrates. Let us begin by examiningthe structureof the kidney, using the mammalian kidney as an example. This will familiarise you with the terminologiesthat are used to describe the anatomicalcomplexityof the vertebrate kidney. When the kidneys fail in a human being, the person can be put on an artificial kidney temporarily. (See Box 9.1) Box 9.1 COIand ur dilyrir lanpcmmb l h didpis fluid fluid In case of renal-failurein humans, the patient has to be put on an artificial kidney. Haemodialysis or CAPD (Continuous- Ambulatory Peritoneal Dialysis)are employed as the techniques. Artificialkidney is a mechanicaldevice which can separate waste products from blood and works on the principle of dialysis. The process employed is called haemodialysis(GK: haeme -blood; lysis- 'separate'). A semipermeablemembrane made of porous cellophane separates large non-diffusible particles like blood cells from small diffisible molecules of urea and other waste products. 9.5.1 Structureof Kid ey P The mammalian kidney (Fig. .9) has a smooth external surface. The typical e metanephric, mammalian kidn y (Refer subsection9.2.2) is a bean:shaped organ attached to the dorsal wall and is retroperitonealin position. It has at its medial side a cavity or depression called hilum or hilus through which the ureter leavesthe kidney. At 'the exit point of the ureter, a renal vein also leaves the kidney and a renal artery and - - nerve enters it.
- 52. FunctionalAnatomy of - Chordates-I1 Fig. 9.9: Sccli)n of mammaliankidneyshowingcortex, medulla and departureof ureter. A section of the kidney revealsthat the kidney is surrounded by a capsule of connective tissue, under which lies the cortex. The renal corpuscles and the convoluted portions of the secretory tubules lie entirely in the cortex (Fig. 9.10). Immediately beneath the cortex is the medulla of the kidney which is striated in appearanceand containsthe loops of Henle and collectingtubules. The medulla is partially composed of large areasknown as medullary pyramid0 or renal pyramids (Fig. 9.9). The medullary pyramids are made up of parallel tubules called medullary rays. Each medullary ray contains one or more collectingtubules and nephrons. Parts of the cortex between the medullary rays form the renal column of Bkrtini. The outer borders of the pyramids are subdivided into smaller units called lobules. The collectingtubules lie in the pyramids but may extend well up towardsthe cortex. The collectingtubules of nephronsjoin together to form collecting ducts or papillary putts of Bellini. There ductswiden near tips of renal or medullary pyramids. The inndr portion of each pyramid is in the form of a blunt papilla and projects into an out pocketihgof the pelvis known as minor calyx (plural: calyces). several minor calycesjoin togetherto enter the major calyx, which in turn opens into the renal pelvis. Urine produced by the kidney entersthe minor and then major calyx. The pelvis leads to the ureter which empties into.thebladder (cloaca in monotremes). Urine which is stored temporarily in the bladder passes to the outsidethrough the urethera. Mammalian kidneys have two types of nephrons and the ratio of each varies depending on the species(Fig. 9.10). One type the cortical nephron, has its glomerulus in the outer kidney cortexwith p tubulethat extendsonly in the outer region of the medulla. Cortical nephrons which prddominate in the human kidqey (90%) have short loops of Henle. The second type have their glomeruli in the deepcortex close to the edge of the kidney medulla and therefore are called juxta medullary nephron. These which predominate (100%) in mammalb of dry habits have long loops of Henle that dip deeply into the rrtedulla. Kidney tubules show'tremendousvariation in vertebrates. Fig. 9.10: A diagrammbtic representation of sectionof mammalian kidney showinga corticalnephron and juxta mddullarynephron.
- 53. Urinary bladder is present in all mammals. It is a muscular sac, derived from the ventral cloaca1wall. The bladder narrows down and connectsto the outside by the urethra. The lower ends of the ureters except in monotremes open directly into the bladder on its dorsal surface. In monotremes they open into the urethra through small papillae near the base of the bladder. At thejunction of bladder and'brethrathere is an abundanceof elastic tissue. Much of the bladder musculaturein the form of bundles continues down to the urethra. Urine flow ceaseswhen the length of the urethra is increased and diameterof lumen is reduced. In males the urethra passes through the penis to open at the tip of that organ through the external urethral orifice or urethral meatus. In femalesthe condition varies. In some, as in rat and mouse the urethra opens independentlyto the outside, passing through the clitoris; in others it enters a urinogenital sinus or vestibule, which is the terminal part of the genital and urinary tract. 9.5.2 Blood Circulation in Kidney Kidneys are highly vascular(Fig. 9.1 1). The renal artery deliversblood to each kidney. Before enteringthe kidney it divides into two branches -one going to anteriorpart of kidney and other to the posterior part. The branchesgive interlobular arterieswhich are located in the renal medullery pyramids. At corticomedullaryjunction, they form the arcuateartery. Fig. 9.1 1 shows the supply of blood to the nephron. Afferent arterioles arise from the interlobulararteries and supply blood to the glomerular capillaries. Blood passes from these capillaries into efferent arterioles. Efferent arteriolesbranch to form peritubular capillary network which nourish PCT and DCT. They also carry away absorbed ions and molecules. The efferent arteriolesassociated withjuxtamedullary nephrons form longthin capillary vessels which run straight into the medulla and loop back towards the corticomedullary boundary. These are called vasa recta or straight vessels. 'They provide nourishment and oxygen to medulla sincethey contain filtered blood. Renal veins take the same course as renal arteries. Blood from interlobular vein form arcuatevessels. The interlobular vessels then converge to form renal vein through which blood leaves the kidney (Fig. 9.11). . Renal Interstitiurn: In both cortex and medulla, the spacesbetween the urinary tubules, contain cells called interstitial cells, which secrete prostaglandin. - . - -- I;ig. 9.1 I:Scctiolb of kidney showing Moodcirculation. SAQ 1 Fill in the blanks with appropriatewords. i) The bench of capillariessurrounded by the Bowman's capsule is called ....................................... ii) A notch present on the medial side of human kidney is called ...................................... iii) .........................artery delivers blood to each kidney. iv) Urinogenital system is ..........................in origin.
- 54. FunctionalAnatomy of Chordates-11 9.6 PHYLOGENYAND SUCCESSIONOF KIDNEYS - - - - - - Comparative studieb of vertebrate development indicate that the ancestral kidney called archinephros(ancieht kidney) of the earliestvertebrates extended along the length of the co6lomic cavity and was made up of segmentally arranged tubules, each resembling an invertebrate nephridium. Each tubule of the archinephros(also called holonephros) opened at one end ihto the coelom by a nephrostome(ciliated funnel) and at the other end into a common nephric duct called archinephricduct. The archinephricduct of each kidney runs along the dorsal side of the coelom and opens into the cloaca. A segmental kidney is found in the embryosof hagfishes and caecilians(Fig. 9.12 a). Kidney of living vdrtebrates have developed from this primitive plan. Duringthe embryonic life of the amniote vertebrates, the threetypes of kidneyspronephros, mesonephros and metanephros (Fig. 9.12) appear in succession. There is a succession of three developmental stages of kidneys, so that this is termed 'succession of kidneys' or 'tripartite concept of kidney' (Fig. 9.12 b). Somebut not all of these Stagesare observed also in other vertebrate groups. F $ . 9.12: Comparatb'e developmentof nialevettebrate kidney. (a) archinephros(b) pronephros(c) mesonephros(d) metanephros. (Light colour representsdegeneratedor underdeveloped structuresand bright red the functionalstructures). Pronephros In all vertebrate embryos, the first kidney to appear is the pronephros. In it the pronephric .tubulesare anteriorly located and segmentally arranged. One end of these tubules open into the coelom y a nephrostome while other end opens into the archinephricduct which in this case is cal 1ed pronephric duct. The two pronephric duct, one on each side extend posteriorly a ~ d *en intothe cloaca. Pronephros is located anteriorly in the body. Only in the adult hagfishes (anamniote) does the pronephros become part of the'persistent kidney also called "head kidneyn (Fig. 9.I2 b). In all other vertebrates it degeneratesduring development an4 is replaced by a more centrally located mesonephros. I
- 55. The term 'opisthonephros" of the embryonic anamniotesis not quite comparableto'the mesonephros of the embryonic amniotes,even though the two are structirally similar. By conventionthe term mesonephros is used for the structure which appears during embryonic development in reptiles, birds and mammals. Mesonephros In the vertebrate embryo, tubules arising from the middle of the nephric ridge constitute the mesonephros (Fig. 9.12 c). Mesonephros is a sometimesknown as the Wolffian body. Mesonephrictubules are segmentallyor metamericallyarranged in the beginning but as more tubules rise this metamerisms is lost. The tubules open into an already existing pronephric duct and no new duct is formed. When the pronephros degenerates the persistent pronephric duct is called mesonephricduct or the wolffian duct. The mesonephricduct may enlarge at its posterior end to form the (i) urinary bladder for storage of urine or (ii) the seminal vesicle for storage of sperms. The mesonephros is the functional kidneys of embryonic amniotes (reptiles birds and mammals) and contributesto the adult kidney (called an opisthonephros) in anamniotes ofjawless vertebrates, fishes and amphibians(Fig. 9.12 and Fig. 9.13). Fig. 9.13: Vertebrate excretory systems (a) comparison of the evoldtion (i-iv) and (b) embryonic development (v-vii) of the vertebrate kidney and its duct. ' Mesonephros may persist after birth in reptiles,the egg laying mammals (Prototheria)and pouched mammals (the metatheria). In the adult amniotes,the functional kidney is the metanephros. As soon as the metanephros becomes functional, Wolffian duct (mesonephric duct) degenerates in females and persists in males as urinogenital duct. llrinogenital System The term archinephricduct is appliedto the kidney duct in anamniotes. The name wolftian duct is given to the duct in amnioteswhich forms in connection with the pronephrosand mesonephros.
- 56. Functional ~natoniy of Chordates - I1 Parts of the male genital ducts such as epididymis and ductus deferens as well as the seminal vesicles develop from mesonephric tubules. Metanephros During embryonic developmentof amniotes-(the reptiles, birds and mammals)the pronephros, mesonephrosand metanephrosappear in succession. The adult kidney of amniotes is the methnephros (Fig. 9.12 d). 'The metanephrosdevelops as the posterior part of the nephric ridge and during development, it gets displaced anteriorly and laterally. The displaced metanephrosdevelops its own new duct, the metanephric duct or the ureter. The dilated tip of metanephricduct becomes the renal pelvis (Fig. 9.9). The metanephrosis is distinguished in several ways from pronephros and mesonephros. It is more caudally (posteriorly) located and is a much larger more compact structure containing a very large number of nephric tubules. It is drained by a new duct, the ureter, which develops when the old archinephricduct is relinquished to the reproductive system of the male for sperm transport. Thus the three successive kidney types -prosnephros, mesonephros,metahephrossucceed each other embryologicallyand to some extent phylogenetically in amniotes(Fig. 9.13). > 9.7 FUNCTIONS OF URINARY SYSTEM The urinary system has several functions (Fig. 9.14): I . The kidneys produce urine which contains metabolic waste products such as ammonia or urea or uric acid etc dependingon the animal group. Urine passes through ureters to the bladder. It is temporarily stored in the bladder and is then released to the outside via urethra through urinary or urinogenital opening. Urine is formed by filteration,reabsorption and secretion (Fig. 9.14 a). W h m r W m n m E o f b l m d h l b a a i n ~ h ~ g u ~hi& nkinblmdmamic pmsmrhub Inmeifid fluidoprorrocproninSpodulmussimulued wur 0' urs b k d N r l t I l q m h h of blmdv * . r mmd b k d p n r m r r . b m o mN m ' radmrglm Rnndnaaltubuk 4 Plnrmmvolume ramred C 4 Blmdprcuurc d ~ z e d Fig. 9.14: Functions oqurinary system (a) removal of metabolic wastes by lilteratius, reabsorption and secretion (uliine formation) (b) osnloregulation (c) regulation of blood volume.
- 57. 2. Kidneys regulate fluid and electroyte balance and hence serve in osmoregulationand water bal-ance(Fig. 9.14 b). 3. Kidneys produce an enzyme renin which has a function in regulating blood pressure (Fig. 9.14 c). 4. The cortical cells of kidneys secrete a hormone Erythropoietin which is associated with production of blood cells. In case of loss of blood or hypoxia, erythropoietin formation is stimulated. Erythropoietinacts on bone marrow to promote formation of erythrocytes. 5. In certain male vertebrates, apart from carrying urine, the ducts from kidneys also carry sperms and are thus called urinogenital ducts. In female vertebrates,ureters act only as a passage for urine. Zlrinogenital System SAQ 2 i) Name the a) Functional unit of the kidney. b) Tuft of capillaries surrounded by renal capsule. c) U-shaped loop between PCT and DCT. ............................................................................................. d) Collecting duct. ............................................................................................. ii) State the parts of the uriniferoustubule in its proper sequence. iii) Tick the appropriate functions of the kidney. a) Removal of metabolic waste. b) Osmoregulation. c) Transmissionof gametes. d) Secretion of renin and erythropoietin iv) Match the following in column A with terms in column B. Column A Column B a) capillaries of efferent arterioles i. Prostaglandin b) Renal interstitium ii. Juxtaglomerularapparatus c) macula densa iii. Red blood cells d) erythroporetin iv. vasarectae v. Sperms 9.8 VARIATIONS IN THE URINARY SYSTEM PLAN You have already learnt in section 9.7 of this unit that the two major functionsof the kidneys are (I) excretion of metabolic wastes and (ii) maintenanceof water and electrolyte balance (osmoregulation). It has been observed that apart from the small variations in structure and function of kidneys that have evolved in the urinogenital systemsof vertebrates due to living in different habitatsthe pattern of the renal system is fairly uniform and apparently simple both in the anamniotesand amniotes. However, study does show that there are great differencesfrom group to group in the construction . of the urinary system in different vertebrate groups these namely Agnatha, Fish, Amphibia, Reptiles and Mammals. This is due to difference in habitat. 9.8.1 Habitat Related StructuralVariations Vertebrates evolved in water. Some emerged on land. Life in freshwaterand in the sea offered different problems of osmotic concentration. Since kidneys were the organs for osmoregulation, the kidney tubules underwent structural modificationsto meet the regulatory needs of each vertebrate group in its own specific environment. Kidneys of vertebrates are confronted with two kinds of problems:
- 58. Functional Anatomy of 1. Water eliminlation in the fresh water organisms and Chordates - I1 2. Water consertvation in the marine and terrestrial organisms. Fresh Water Organism In.freshwater organisms,kidneys have (i) large well developed glomeruli which produce large volumes of glbmerular filterate. (ii) Prominent distal tubules absorb salts and amino acids from the filtrateto retain them within the body and also absorb much less water. Lots of water is eliminated in the urine. . Marine Organism Marine organisms face dehydrationas the outside environment(sea water) has greater osmotic concentration.So water tends to move out of the body of the marine organism. The structural modificationsin these organisms is adapted to retain water and are: i) Loss of glomeruli (aglomerularkidney). ii) Shorteningor loss of distal segments of kidney tubules which are responsible for reabsorption of salts. Both the above modificationscause water retention and increased salt excretion so that homeostasis is maintained in these organisms. iii) Marine fishes have extra renal structures for excretion of salts. Elasmorbranchs (cartilaginousfishes)have chloride secreting glands on the surface of gills and also rectal glands which secretesalts. Marine reptiles and birds have salt excreting- paired large nasal glands located in a bony socket with duct opening into nostril. Refer to unit 9 of LSE-05. Terrestrial Organism Terrestrialorganismsalso need to conservewater. Snakes have aglomerular kidneys for this purpose. In mammals, the Henle's loop of the nephron concentrates the urine by reabsorbingwater firom it. The length of loop of Henle's varies with the environment of the organism. For axample beaver which have lots of water in their immediate environment have short, Henle's loops. Both long and short loops of Henle are present in rabbit and humans which have an intermediatecapacity for concentrating urine (see Fig. 9.6 again). The sand rat, on the other hand which is found in arid conditions has very long loops of Henle's in order to reabsorb as much water as possible and so excretes a very concentratedurine. 9.8.2 Variatians in Urinary Systems of Vertebrates belonging to Differeat Groups The vertebrate kiddeysthemselvesare varied in structure; the ducts vary as does the urinary bladder. These variation are due to two major causes: (I) The kidney unlike many organs must start functioningat an early stage of development in order to remove the metabolic wastes of the rapidly growing embryo. The kidney is however, subjectto modification or replacement in the later developmental stages and adult existence (ii) as the gonads (testis and ovary) lie adjacent to the kidneys. These organs specially the testis tend to take over part of the tubes and tubules of the urinary structures as a conducting system for their prdducts. Asa result of this the urinary organs have been markedly modified in most vertebrategrcups. In this subsectionwe will survey the construction and hence normal variationsof urinogenital systems in various vertebrate groups (Fig. 9.15 a tog). Agnatha (Cyclostome) or Jawless Vertebrates The adult kidney ofjawless vertebrases lamprey (Petryomyzon) is opisthonephros or Mesonephronsand the tubules are derived from the posterior mesomere. The tubules have lost all connection with the coelomic cavity but form a close association with the glomerular blood vessels at the blind-ended kidney capsple. The absence of connection between kidney tubules and coelom makes it evident that "coelomic fluid filter" was replaced by "blood filter system" early in vertebrate evolution. This may be because possibly coelomicfluid pressure is too low and variable to keep up with the fluid balance demands of vertebrtates which have high metabolic rates. In adult lamprey the kidney is elongated and flauened. The kidneys lie on either side of the mid-dorsal line and each is suspended by a mesentery like membrane. The
- 59. archinephricduct courses along the free edge of the kidney. Tlie pronepliros exists above the ~nesonepllros as the non functional "head kidney". In the adult hagfish the opisthonephros consists simply of a series of tubule arranged in a segmental manner along most of the length of the trunk, each tubule draining directly into the archinephric duct or pronephros. In hagfish adults the archinephrosbecomes modified to form a persistent head kidney. The remainder of the kidney, however is separated from the head kidney and becomes the opisthonephros. The hagfish has only forty glomeruli, each connected to the collecting duct (archinephricduct) by a s~nall neck segment. (Fig. 9.15 a) Fish The kidneys'of fislies are opisthonephricor mesonephricand though tliey exhibit great variation in shape, they are however funda~nentally similar in structure(Fig. 9.15 b and c). In all species they are dorsal in position. In some speciesthey extend almost the entire length of coelom. In other fishes tliey may be more voluminousand the two sides may sliow various degree of fiision. In still otliers they are sllort and confined to the posterior part of the body cavity. Peritoneal funnels are only retained in a few forms, notably Anria, Sturgeons and certain elasmobranchs. In some marine teleosts no external or internal glomeruli are present, and so such kidneys are called as aglomerular kidneys. Generally the kidneys of male fisli are longer than those of females, sincethe anterior ends in male are appropriated by the reproductivesystem. In males of some groups small modified kidney tubqles now called efferent ductules,connect the testes with tlie archinepliricduct. This archinephric duct is termed as the ductus deferens and serves as the passage for sperm transport. It may, in addition continueto carry wastes. However, in such cases there is a marked tendency for the posterior portion of the opisthonephrosto take over the greater part of the excretory function, with one or more accessory ducts developing which are responsible for carrying wastes directly to the cloaca or to the outside. In Selacliians,Chondrosteans and some otliers the conncectionof the testis and archinephric duct usually occurs at the anterior end of tlie ~pisthonephros. In teleosts however the testes and opisthoneplirickidneys are not connected. In them the duct from the testes eitherjoin the archinephricducts near the posterior ends or open independently to the exterior. In some fislies the dilation of the archinephricduct may form a bladder like enlargement for temporary storage of urine (Fig. 9.15 c). In those fish where archinephricduct functions as ductus deferens, enlargements called seminal vesicle and sperm sac may develop to serve as temporary storage places for spermatozoa, Amphibians Tlie amphibian kidney is opisthonepllricor mesonephric and the anterior tubules ofitlie opisthoneplirosfunctions as tlie sperm duct. The primitive archinephrictype of kidney is found in tlle larval caecilians of the amphibians. The kidney is similarto the larval kidney of the hagfish and consists of the distinct ~netameric arrangementof kidney tubules, renal corpusclesand nephrostome. In the adult the kidney is opisthonephros.The opisthoneplirosis lobulated and extends the greater part of the length of the coelom. In many larval amphibians a small head kidney with peritoneal connection may be present but it does not persist in the adult. The urodele amphibians have opisthonephricor mesonephric kidneys that are similar to the elamobranchs (cartilaginous fisli). The kidneys have two regions (i) anterior narrow region which in males is more genital than urinary in function and is referred to as epididymis and (ii) posterior expanded portion which is the main part of the opistho~iephros and is called the 'kidney proper'. The archinepliric ducts run along the lateral edge of tlie kidney a short distance from tlie kidney proper. Numerous collecting ducts or tubules which are more developed in males than fe~iiales join at intervals to tlie archinephric duct from the opisthonephros.The archinephric duct now called mesonephric ducts serves in tlie male as a ductus deferens as well as transportswastes but in females is concerned only with transporting wastes. The mesonephric ducts in both sexes open into the cloaca on either side. through a small papillia (Fig. 9.15 d). llrinogenital System
- 60. In the anurans the opisthonephrickidneys are dorsoventrally flattened have a more posterior concentrationof tubules. They are confined to the posterior part of the abdominal cavity and so are retroperitoneal and dorsally located. Unlike in the urodeles the anterior and posterior regions of the kidney are not clearly distinguished.An yellowish, orange adrehal gland is closely attached to the ventral side of the kidney. In femalesthe kidneys anid reproductivesystem have no connectionto each other. However, in males they are intimlatelyconnected. In males certain anterior kidney tubules become modified into efferent ductulesthat connectthe testis to the kidney while the mesonephric duct servesto transport spermatozoaas well as urinary wastes. In these animals unlike the urodeles the archinephtic duct is located within the kidney along its lateral margin. It leaves the opisthonephfosnear the posterior end and passes to the cloaca (Fig. 9.15 e). Openin8ofu m e r intoclorr Bird(Aves) Fig. 9.15: Urinary systemsof vertebrates(a) cyclostome(b) cartilaginousflsh (c) bony fish (d) salamander (amphibian), (e) frog (amphibian), (f) lizard (g) bird.
- 61. A thin walled, bilobed bladder which is thought to be endodermal in origin and homologous with the urinary bladder is present. It is derived from the cloaca1wall and opens into the amphibian cloaca a short distance beyond the openings af the aichinephric ducts. The archinephric duct and bladder are not connected directly, and so the thin, watery urine tirst passes directly into the cloaca. Reptiles Reptiles are the tirst co~npletely terrestrial vertebrates adapted to fresh water, estuarine and even marine habitats. Kidney in reptiles are metanephric. In reptiles the kidneys are usually small and compact with lobulated surface arid confined to the posterior half of the abdominal cavity, generally to the pelvic region (Fig. 9.15 f). The posterior part of the kidney narrows down on each side and in some lizards the hind parts may even fuse. The degree of symmetry varies in reptiles especially in case of snakes and limbless lizards who due to their elongated body shape possess excessively lobulated, long, narrow kidneys; often one kidney may be entirely posterior to the other. In snakes and crocodilians a urinary bladder is absent. Most turtles and lizards however have well-developed and usually bilobed bladders which open into the cloaca. Except in turtles the ureters open separately in the cloaca. In turtles the ureters are connected to the bladder. In some turtles a pair of accessory urinary bladders is also connected with the cloaca. These functions as accessory organs of respiration. In females they may be filled with water. which is used to soften the ground when they prepare their nest. Living reptiles have only about a few thousand nephrons in which loops of Henle are absent and renal corpuscle are poorly developed. The glomeruli are small in order to conserve water. Birds The kidneys in all birds are situated in the pelvic region ofthe body cavity and their posterior ends are frequently united. They are complex, lobed structures with short ureters, which open independently into the cloaca (Fig. 9.15 g). The kidneys contain some mammalian-like kidney tubules with loops of Henle (some short and some long loops) that parallel collecting ducts. Birds mostly, however have reptilian type tubules that do not have loops. Urinary bladder is absent in all birds except for ostrich. Urinary wastes are ~nainly in the form of uric acid and are eliminated via the cloaca along with the faeces. The ability of l<idneyto concentrate urine and thereby conserve water is not as good as in mammals. but is better than in reptiles. The cloaca and even the posterior large intestine may further modify and concentrats urine by resorption of water and ions. The mammalian urinary system has already been dealt is the previous sections. Refer subsection 9.2.2, sections 9.4, 9.5 and fig. 9.4 for details. SAQ 3 i) Tick the correct alternative: a) Marine organisms possess I) sI101-tdistal tubule 2) long distal tubule 3) very long distal tubule 4) short proximal tubule. b) Snakes have aglomerular kidney in order: I ) to conserve water 2) to remove water 3) to conserve protein 4) to remove protein. c) The desert kangaroo rat in order to combat dehydration : I ) has long Hen le's loop for reabsorption
- 62. 2) utilises metabolic water 3) eats waterladen vegetation 4) all of the above. - 9.9 THE GENITAL SYSTEM The genital system is tlie reproductive systetn and is made up of tlie gonads, gonoducts and genital openings. The gonads as you have read are mesodermal in origin. They are called testes (sing testis) in males and ovaries in female. Both the testes and tlie ovaries produce gametes and secrete hormones and are termed primary sex organs. The gonads are usdally paired structure, though unpaired gonads occurs in some forms such as cyclostomds aawless fish), certain fish as well as female birds of most species. This is the result oteither fusion of paired gonads or the unilateral degeneration of one gonad. Evidence of metamerism of gonads in chordates is only found is the primitive ampliioxus - cephdochordate. You will recall that ann~nniolcs include In vertebrates the dvaries and testes come to be attached to the dorsal body wall by cartilagillousand bony tishes and mesentery like bantls of tissue, the mesorchium in the male and mesovarium in the amphibianswhile amniotes include the fernale. In most ve~ebrates the gametes (ova in females and spermatozoa in males) reptiles, birds and mammals. produced by the gdnads are transported outside the body by means of the deferens ducts (vas deferensb in males and by oviducts in females. In a few forms like cyclostonie ducts ake absent in both sexes. Eggs and sperm escape from the body cavity through genital pores. The deferent ducts as you will recall are usually the mesonephric or tlie Wolffian ducts which also serve to transport urinary wastes from the opisthonephric ior mesonephric kidneys in those animals in which these kidneys function either during embryonic or adult life (in male anaminotes). In arnniotes in which the metanepliros is tlie functional kidney and in which the mesonephros degenerates, the Wolffian duct of the male on each side persists to become the ductus deferens (male genital duct). In most vertebrates in both sexes when the reproductive ducts first develop they open posteriorly into the cloaca. This relationship persists throughout life in many vertebrates, but in others tlie cloaca1 region becomes modified so that the reproductive ducts either open separately to the outside or in the males atleast, join the excretory ducts to emerge by a common opening or urinogenital opening. In many aquatic vertebrates fertilisation is external while in all terrestrial vertebrates except for anurari amphibians and even in many aquatic species it is internal. In some animals transport of spermetazoa from male to female is brought about by apposition of the cloaca (c1oaca:sing) of the two sexes in most animals, however, males have copulatory organs which are used in an intromittent manner to deposit the spermatazoa into the reproductive tract of the female. Various types of copulatory organs are found in the vertebrate groups. In both sexes all the structures or organs which help to bring the germ cells or products of the primary sex organs together are termed as accessory sex organs. These include the * reproductive ducts, associated glands and intromittent organs. Secondary sex characters are indirectly concarned with sex but play a part in the reproductive scheme. Sexual differences in such secondary sex characters like plumage, body size and strength, as well as vocal apparatus are only indirectly related to reproduction. Cloaca (Roman term for sewer) is present in a variety of vertebrates. It occurs as a ventral pocket, at tlie back end of the trunk of the animal, and opens to the exterior. The orifices of tlie digestive, genital and urinary systems open into it. The cloaca appears to be a primitive vertebrate feature. Primitively, tlie gut. urinary duct and genital ducts all terminate in a common, short chamber, the cloaca that empties to the outside. Most tetrapods (amphibians, birds reptiles, egg laying mammals) have retained a distinct cloaca, although tnost mammals have not.
- 63. SAQ 4 Fill in the blanks witli appropriate words. i) The primary sex organs are the ...........................and ................................ ii) Mammary glands are .................................... sex organs. iii) ...................................... duct becomes the ductus deferens. iv) The ................................. are attached to the dorsal body wall by the mesovariu~n. ......... v) When reproductive ducts first develop, they open into ........................... vi) The tissue that s~~spends the testis from tlie dorsal body wall is called .................................... vii) Copulatory organ is an ....................................organ. 9.9.1 Embryonic Origin of Gonads and Gametes The sex of an individual depends basically upon the nature of its chromosomal inheritance. The early development of tlie embryo is mainly because of the organisation already present in tlie unfertilised egg, the influence of the sperm and the hereditary features it introduces, all which are not obvious until a relatively late stage. We thus observe that for some time the sex organs of tlie embryo remain in an indifferent stage during which the dcvelopmcnt of the gonads and their ducts proceed considerably far without any indication of whether tlie embryo would develop into a male or female. Finally the embryo develops its specific sex features, presumably due to hormonal activity. Embryonic origin of Gonads Maleness in chordates develop basically on a female plan. Both the gonads and kidneys develop from tlie intermediate mesoderm (mesomere) of the embryo (Fig. 9.16 a). In vertebrates there is a great difference in the rapidity with which different organ systems develop. The nervous system for instance, grows very rapidly in early stages while the genital organs are one of the slowest to develop. The gonads make their appearance only at a stage when most of the other organ systems have been blocked out and the coelolilic cavities are well developed. Paired longitudinal swellings the genital ridges develop along the roof of tlie coelom, medial to the embryonic kidney and on either side of the dorsal mesentery (Fig. 9.16 b). These ridges gives rise to the gonads. The gonads developing from such ridges are elongate to begin with but often in later stages become short and compact with a L I S L I ~ ~ tendency for anterior concentration of tissue. The germinal epithelium of the ridges are continuous witli the mesodermal lining of the rest of the coelom and form the more important structural parts of the gonad. Mesenchyma lying beneath the epithelium forms the connective tissue and in higher vertebrates at least gives rise lo tlie special interstitial tissues that are thought to be a source of gonad hormone. Before the end of the indifferent stage, the gonad generally develops in many cases into a swollen structure which extends out into the coelomic cavity from its dorsal wall in the neigbourhood of the developing kidney and is often supported by a special mesentry. From the germinal epithelium that cover the gonad surface, finger like structures called the primary sex cords grows inward into the underlying mesencliyma of the gonad (Fig. 9.16 b). The gonads of both sexes initially contain germ cells. These germ cells do not arise in the genital ridge nor even in the ad-jacent mesoderm. In fact they do not arise in the embryo at all. They f?rst rise in remote sites outside the embryo in the extra-embryonic endoderm. From here they migrate to tlie indifferent gonad where they get located permanently. In females the germ cells establish residence in the cortex while in males they get located in the medulla. In ;impliibian males. 1l1ecloaca1 glands are prescnl ,liicli secrele scell1 in order lo allracl the female for mating. In them clocal glands also secrete ;I jelly like substance vhicli holds logether a package of sperlns which is called spcrn~rtophore. The l'emale picks up the spermrilopliore in her cloaca for fcr~ilisatio~l of her eggs. As gonads mature they enlarge and are pushed downwards where they lie suspended by a dorsal mesentry termed mesorchium in males and mesovarium in females.
- 64. Functional Anatomy of Chordates - 11 ~ p l n s l LO^ --& Glomerular capsule - L x Embryo cross section Glomerular capsul c Porm~tion of female reproductive system Van defdrens Urinarybladder i Proslalc U Formution of malc re()rsductive syrtem Fig. 9.16: Developmentofgonads in vertebrates showing modification of the indifferent gonadinto ovary or testes. Thegonadal structure, whether primary cortex or medulla is derived from embryonic mesoderm(a) the primodial germ cells which give rise to spermatozoaor egg are initially located in the beginning, within the embryonic endoderm and then migrate through the mesenteries during development into the indifferent gonad (b) the germ cells arrange themselves betweenthe medullary and cortical cells of the ind~fferent gonad (c) the cortical tissues predominatein the formation of the ovary while (d) the medullary tissues predominates in the for motion of testes. 9.9.2 Functions of the Genital System The primary function of the genital system is production of gametes, the sperms and the eggs respectively by the male and female gonads under the influence of hypothalamol-pituitary hormones. Gonoducts of the genital system transmit gametes to the place most suitable for fertilisation. It may be water in which eggs are laid by the female and sperms dropped on them by the male (external fertilisation) or sperms may be discharged just as far into the female tract where egg and sperm can meet (internal fertilisation). Transport of sperms into female body is specially performed by accessory reproductive organs or intromittent organs, to ensure fertilisation. The female gonoduct is different from that of the male. It is specialised for retention of: (i) egg (in case of egg laying or oviparous organisms), e.g. fish, amphibia, reptiles and binds (ii) the fertilised egg with the developing embryo (in case of ovovivipardus organisms which lay the egg after embryo has developed to a cemin stage) e.g. certain snakes or (iii) the developing embryo (in case of viviparous organisms which deliver full fledged young ones) e.g. mammals.
- 65. llrinogenital System Gonads secrete sex hormones which are steriod in chemical nature. The male hormone secreted by the testis is testosterone. The female hormones are malnly estrogen and progesterone. The sex hormones control the production of secondary sex characters, difference in structure, physiology and behaviour by which male and female can be distinguished. The secondary sex characters are especially prominent during breeding season. SAQ 5 Choose the correct alternatives from the parenthesis. i) Reproductive organs develop from the embryonic (mesodennleaoderm) germ layer. ii) The finger like process developing from germinal epithelium are called (primarylsecondary) sex cords. iii) Organism that lay eggs in which the embryo develops are called (viviparous1oviparous). iv) Mammals are (ovoviviparous1viviparous). v) Estrogen is a (malelfemale) hormone. 9.9.3 Genital System of Protochordates Branchiostorna is a dioecious chordate. The gonads of both male and female are similar in arrangement, with distinctly segmented gonads (i.e. is many gonads) that are arranged below the muscle segmenrs or myomeres (Fig. 9.17 a). Approximately 26 pairs of gonads on either side of the atrium also called peribranchial space project into it from the inner surface of the body wall. The most anterior gonads are located around the middle of the pharyngeal region. Each gonad (or segment) is a hollow sac which is lined with an (I) outer coelomic epithelium and an (2) inner germinal epithelium. Gametes develop within the cavity of the gonad and are expelled through the wall into the artrial cavity by means of temporary pores. From this cavity the gametes pass directly to the outside through the atriopore. In these animals fertilisation is external. Overall, the reproductive organs of the Branchiostonta are not similar to those of vertebrates, The ovarian follicle also appears to differ from the vertebrates because a follicular epithelium is absent. Urochordates (Tunicates) are he~maphrodites, that is, they have both sex organs in the same individual. In other words they are hermaphrodite. Ducts from gonads enter the atrium, a space surrounding the viscera (organs of the body) and then to the outside by the atriopore (Fig. 9.17 b). I*ww~ n11 I ~ U I hwnof smvr Ntwlurrd Ihwnl nonu IXnsulruhmiewnal t;cladrmral rfilhcrllum Pnnr?nr tnmrrerrr mu.** I t ~ b n n n A l f N n Fig. 0.17: Genital systclns ol' protochordates (a) cross section ol'Brmrrltic~stomo,showing segmental gonads (b) saggital section of a tunicste showing its reproductive systems. I . Paired gonads -testes (singular - testis) 2. Paired urinogenital ducts
- 66. I Functional Anatomy of Chordates - 11 3. Single urinogenital opening Let us begin our studyof the male genital system with the anatomy of the testes. 9.10.1 Testes Testes of all vertebrates have similar construction. Thetypical testis is a compact organ whose shape however varies in members of different vertebrate classes. In some forms, testis is composed of: (i) elongated tubules called the seminiferous tubules in which the primordial gem ce~lls develop into maturesperm and which connect by means of ducts to the outsidee.g. anulran amphibiansand amniotes.(ii) in others,the testes consists of rounded cavities called seminiferousampullaeor spermaticampullae or spermatic cysts - e.g. cyclostomes,fish and urodeles. Both the rounded ampullae and elongated tubules, at first consist of solid masses of cellswhich later developthe cavities or lumina. SeminiferousTubules The testes in anurans, amniotes(reptiles, birds and mammals) and even some teleosts, are composed largely ofiseminiferoustubules which are coiled tubes and whose walls contain cells that produce sperm (Fig. 9.18 a). I Fig. 9.18: nlwmmnlian testis (a) section through testis show~ng spcrm passage (b) nn enlargement of the boxed nrer in (a) showing the details of seminiferous tubules and rete testis (c) section of kidne) showing seminiferoldstubules and a group of Leydig cells between the tubules (d) enlargement of the boxed area inkc) showingdcvcopingsperms and sertoli cell.
- 67. The testes are surrounded by a capsule, the tunica albuginae. Seminiferoustubules may constitute upto 90 per cent of the testis. The tubule walls consistof a multilayered germinal epithelium containing spermatogoniccells and Sertoli cells which are nutritive in function and in which the heads of the maturing spermsare embedded. Sertolicells probably also produce, most of the fluid in which the sperms are suspended,while leaving the testis by means of active filteration from blood plasma (Fig. 9.17 b). Seminiferoustubules may end blind at the tunic or outermosttissue layer, and pass toward the centre, becoming tortuous, that is, full of twists and bends, before emptying into a system of collecting tubule, the rete testis. Such an arrangement is characteristicof frogs. In certain amniotes like the rat for examplethe tubules may be open ended, running a zig-zag course from the rete to the periphery and back again. The average length of such tubules is 30 cms, and they seldom communicatewith each other. In many mammals, the tubules are grouped into lobules separated by connective-tissuesepta, or walls. This arrangement allows the packing in of an extensive quantity of germinal epithelium into a small space (Fig. 9.18 a). The tubules are inconspicuousand epithelium is inactive in immature males and between breeding season in the adult males of those specieswhich have specific breeding seasons. In some species however, spermatogenesisor production of sperms in adult males proceed at a variable pace throughout the year. An active epithelium may exhibit all stages of developing sperms. The lumen or tubule cavity containsthe tails of many spermswhose heads are embedded in thc Sertolicells, free spermsand fluid that is resorbed. In mammals in any single zone along a tubule, all sperm are at the same stage of maturation;adjacent zones contain different generation of sperms and a period of sperm formation and discharge is followed by interval of inactivity. Seminiferous Ampullae Cyclostomes,most fishes and tailed amphibian have seminiferouscysts also called spermatogonial cysts, spermatocysts or sperm folliclesor ampullae or crypts or sacs acini or capsules in which sperms develop but in which the epithelium is not germinal. Sperms mature within the ampullae, among cells called the Sertoli cells. The Sertolicells appear to be partially nutritive in function. Once the sperms mature, the walls of the ampullae break down and release the sperm into the coelomiccavity. The arrangementof the germinal epithelium is different from that of seminiferoustubules. Sperlnatogenic cells migrate into the cysts from a permanent germinal layer, which dependins on the species may be among cysts at the periphery of the testes or in a ridge along one margin of the testis. The spermatogenic cells after invading the thin, non- germinal epithelium of the cells, multiply producing a large number of sperms. The cysts become swollen and whitish in colour and the entire testis swellsup as well becoming granular in appearance. When sperms mature they separate from the epithelium and move freely in the cystic fluid. Finally the cysts burst and the sperms are shed into the duct. In the case of cyclostomesand a few teleosts the sperms are released into the coelom. The cysts collapse on becoming totally empty. They are then either replaced by new ones or become repopulated by additional spermatogeniccells. The spaces between the seminiferoustubules or spermatogeniccysts in the testes are filled with testicular stroma which consist chiefly of connectivetissue, blood, lymphatic vessels and nerves. Stroma is more abundant in some vertebratesthan in other. Glandular interstitial cells called Leydig cells are also present in most if not all vertebrates. The Leydig cells are thought to be a primary source of androgens or male hormones. These cells are not always easily distinguishable.The capillar%wystemof the rat testis and probably that of many other vertebrates is such that blood which bathes the Leydig cells flowsto the tubules. In most vertebrates however the adult gonads retain a position in the upper part of the coelomic cavity. In vertebratesexcept mammal the testis lie within the body. This is also the case in many and sometimes in all members of the mammalian orders Monotremata, Insectivora, Hyracoidea,Edentata, Sirenia,Cetaceaeand Proboscidiea. In some male mammals like in most marsupials, ungulates, carnivoresand primates after infancy the testes descend into special pouch called scrotum where they are lodged permanently. The scrotum or scrotal sac is a temperature regulatingdevice. The scrotal sacs are paired structures which project externally from the floor of the abdominal cavity, each connected to the abdominal cavity by a inguinal canal lined with peritoneal membrane. During development the testis move backward and downward from their Ilrinogenital System
- 68. Functional Anatomy of Chordates -11 original position into these sacs each accompanied by its duct, blood vessels, lymphatic vessels and by a fold of its proper mesentery called gubernaculum. A few mammals have a pouch into which the testes descend and from which they can be retracted by the action of the musclie (cremaster) in the scrotal sac. ' Ihese include a few rodents such as ground squirrels, most bats and some primitive primates (loris, potto). In some cases the sacs remain in open connection with the abdominal cavity, and the testes may be withdrawn into the body between breeding season through the iguinal canal which acts as path of descent and retraction of the testes to the sac. In descending, the testes carry along a spermatic duct, blood and lymphatic vessels and a nerve supply wrapped in peritoneum which collectivelyconstitutethe spermatic cords. In rabbits, most rodents and some insectivores the scrotal sacs are absent instead they have a wide inguinal canal into which the testes may be withdrawn and from which they are retracted when in danger of injury. In these mammals, the descended testes cause a temporary bulge in the perineal region (i.e. between the anus and urinogenital opening). In a small number of mammals the testes permanently occupy the perineal location. In some mammals including man (Fig. 9.4 a) the sacs may be permanently closed off, but sometimes a weak spot in the abdominal wall of this region may rupture leading to a condition known as iguinal hernia. 9.10.2 Male Genital Duct The male gonoducts or genital ducts which in most vertebrates serve to transport spermatozoa to the outside of the body are the archinephric ducts or the Wolffian ducts, that are formed in connection with the developing kidneys. You will recall that the name archinephricduct is used for the kidney duct in anamniotes. Although the male genital ducts are similarto the female genital ducts, the male genital ducts have a more complex history and organisation. The original function of these ducts as you will recall, is elimination of urinary wastes. In a number of fishes ~ n d amphibianscertain modified kidney tubules are employed in carrying spermatozoafrom the testis to the archinephric duct. 'They are known as efferent ductules and the archinephric duct is then termed as the ductus deferens. Even in the amniotes in which the mesonephros degenerates, its duct persists to become the ductus deferens and epididymis (ductuh epididymidis). This ductus epididymidisestablishes connectionswith the testis via effh-ent ducts or ductules (Refer Fig. 9.18 a) which in this case are the modified and persistent mesonephrictubules. Reproductiveducts are lacking in amphioxlus and modem jawless fish, the cyclostomes. In modernjawless fish the testicular ampullae rupture to release sperm into the coelom from where they pass out of the body by means of the genital pores (Fig. 9.19). In most vertebrates however the sperms never enter the coelom but go directly into genital ducts. In all jawed vertebratesthe testes arejoined to one or more ducts called the sperm ducts through which the sperm leave the body All sperm ducts includeparts of the kidney ducts and so they are called urinogenital ducts. The exception are the teleost fish in which the ducts are not derived from the kidney but from the testis itself. In most vertebrates the sperms mature as they pass through the genital ducts but in teleosts they seem to be fully mature at the time they leave the testis. The genital ducts terminate at the urinogenital sinus which often empties into the cloaca. Uninogenital ducts may also carry urine from the adjoining kidney though that part of the kidney stops functioning in some species. From the testis is formed a tubular network called the rete tubule or rete testis. The rete testis within the testis forms a network of thin walled ductules or minute ducts that collect sperms from the seminiferoustubules (Fig. 9.18 a). The rete is drained by a number of small ducts called the efferent ducts or vasa efferentia. The vasa efferentia are usually modified kidney tubules and are usually less than 10 in number. The kidney tubules or vasa efferentia drain into the archinephric or mesonephricduct aalled the deferent duct in male which empties into the urinogenital sinus. The efferent and deferent ducts are collectivelytermed as epididymis and may be used primarily for sperm transport (Fig. 9.18 a). The epididymisof amniotes which is a highly coiled duct that drains the vasa efferentia usually serves as a temporary storage place for sperm. In mammals the first part of the epididymisconsists of a head, body and tail that wrap around the testis and then gradually straightensout to become the spermaticduct. The epididymis secretes substancesthat prolong the life of the stored sperm and increase their capacity for motility.
- 69. 9.10.3 Male Accessory Sex Glands Vertebrates males, especially of terrestrial species may have several accessory sex glands. Some amphibians (salamanders) have cloacal and pelvic glands which secrete ajelly package to enclose sperm thus forming the spermatophore.They may also have other functionsas well. In mammals a number of relatively large and complex accessory glands occur. The testis and epididymis secrete fluids that form part of the semen. Other glands are the prost ate gland. vesicular glands, bulbourethral glands, urethral glands (or Littre's gland) and coagulating glands. All of these open into the urethral part of the urinogenital sinus. Not all the glands are present in every species. However, the major mammalian sex glands are the prostrate, the bulbourethral and the ampullary glands and the seminal vesicles all of which are the outgrowths of the spermaticduct or of the urethra and all four occur in elephants and horses and in most moles, bats, rodents, rabbits, cattle and primates. The prost ate is the ~iiost widely distributed mammalian accessory sex gland. It empties into the urethra by multiple ducts. Many rodents, insectivores and lagomorphs have three separate prostatic lobes. In a few mammals which include some carnivoresand primates, the prostrate is a single mass with lobules and encirclesthe urethra at the base of tlie bladder. In many rodents as well as in some other mammals, the semen coagulates quickly after ejaculation due to secretion from a male coagulatinggland which is usually considered part of the prostatic mass. Coagulated semen forms a vaginal plug that temporarily prevents copulation. UrinogcnitwlSystcn~ Balbourethralor Cowper's glands arise from the urethra near the penis and are surrounded by the muscle of tlie urethra or penis. Usually there is one pair of Cowper's gland in the mammals, except for in some marsupials where as many as 3 pairs have been - found. An ampullary swelling on the spermatic duct near the urethra is present in many mammals. However in a small number of mammals a separateampullary gland is formed as an outgrowth of the duct. Seminal vesicles are paired, typically elongated and coiled fibromuscularsacs that empty into either the spermatic duct or the urethra. These vesicles contributethe sugar fructose and citric acid to the semen but do not serve as sperm reservoir. 9.10.4 Intromittent Organs In most aquatic forms external fertilisation takes place and water providesthe medium by which the spermatozoa reach the eggs. In terrestrial forms, however, a liquid environment is needed to transport the spermatozoansso internal fertilisation is the rule. The necessary fluids are produced by both male and female. In a number of terrestrial vertebrates spermatozoaare transferred from male to female by cloacal apposition but in most terrestrial forms, and even in many aquatic specieswhere fertilisation is internal the males develop.organs called intromittent or copulatory organs which are used by them during internal fertiliation to introduce sperms suspended in seminal fluid into the female tract. A number of types of copulatory organs exists among vertebrates all of which are not homologous. lntromittent organ in Anamniotes . Fish -Among fishes copulation with internal fertilisation occurs only in elasmobranchs, holocephaliansand some teleosts. In elasmobranchscopulation is accomplished by means of clasping organs called claspers (Fig. 9.19 a) which are modificationsof the medial portions of the pelvic fins of males. During copulation one clasper is inserted into the female cloaca. Sperms leave the male cloaca, enter a groove on the clasper and are flushed by water squirted from siphon sacs within the body wall of the male into the female cloaca. In the teleosts which have internal fertilisation the anterior border of the anal fin of the male may be elongated posteriorly to form an intromittent organ called the gonopodium (Fig. 9.19 b).
- 70. FunctionalAnatomy of Chordates - II Fig. 9.19: Intromittent organs of Ashes (a) clasper, u modified pelvic fin of male elasmobranch Squalus acanthias(dbgfish) and (b) gonopodium,which isa modification of the anterior part ofthe anal fin in males, of those teleosts. who exhibit internal fertilisation. Amphibians -Copulatoryorgans are absent in urodeles and anurans though internal fertilisation does oacur in urodeles. In these animals the male deposits spermatophores, which are actually small packets of spermatozoaheld together by secretions of cloacal glands. The femalepicks up the spermatophoreby the muscular movementsof the cloacal lips. A dorsal diverticulum of the cloaca, the spermatheca serves for storage of the spermatozoawhich are thus available for fertilisingthe ova as they pass down the oviducts to the cloaca. lntromittent organs of Amniotes lntromittentorgans are very well developed in reptiles and mammals and few birds like the ostrich, drakes and ganders. In the amniotes intromittentorgans are of two kinds: (1) Hemipenes are paired, saclike organs devoid of erectile tissues lying in pockets under the skin near the cloaca. Hemipenes can be everted or retracted eg. Snakes and lizards (Fig. 9.20). (2) Penis is an unpaired erectileorgan, eg. male turtles, crocodiles, few birds and mammals (Fig. 9.21 and 9.22). Reptiles: The only reptile lacking copulatoryorgans is Sphenodon. In other reptiles as you have learnt, two types of structuresoccur. In snakes lizardsthe hemipenes is present. They are everted during copulation by propulsorand retractor muscles and filling of blood sinuses in the hemipenes(Fig. 9.20). In turtles and crocodiles,the single penis is derived from paired thickenings or ridges in the anterior and venltral walls of the cloaca and is composed of connective and erectile tissues. The paired masses of erectiletissue are called corpora cavernosa. A groove along the dorsal surfaceserves as a passage for the spermatozoa. During the mating act, the corpora cavernoha are filled and distendedwith blood, making the penis firm and enlarged and erect. The penis can be extruded and retracted. Fig. 9.20: lleniipenes of nlrlc reptile (lizard). Birds: A penis occurs only in the males of ducks, geese, swans and ostriches. It is a single structure, buillt on the same plan as'that of turtles and crocodilians. In the remaining birds sperms are transmitted through cloaca (Refer unit 16 of LSE-10). Mammals: A single penis is typical of mammals. In monotremes, under normal conditions,the penis lies in the cloacal floor. It is similar to the organs of turtles, crocodiliansand bird except that the groove on the dorsal side becomes a closed tube.
- 71. In addition the tube is surrounded by a singlemass of erectile tissue the corpus spongiosumwhich is separate from another pair of erectiletissue mass called the corpora cavernosa. The canal in monotremes is supposed to carry only spermatozoa since the urethra has a separate opening into the cloaca (Fig. 9.21). Fig. 9.21: Diagrammatic longitudinal section through cloacal region of male monotreme showing a retracted penis. In the rest of the male mammals the openings are not separate(Fig. 9.22). The urethra in these animals and subsequent groups serves as a passage for both urine and seminal fluid. The two corpora cavernosa are separated by a septum and the corpus spongiosum surroundsthe urethra. The end or tip of the penis is enlarged to form a sensitiveswollen glass, containing erectile tissue and numerous nerve endingswhich make it extremely sensitiveto certain stimuli. It is is continuous with the corpus spongiosum. The glans is covered by a thin and delicate skin called foreskin or prepuce. Fig. 9.22: Sag$:.-.! oicw of pelvis of human male showingthe urinogenital system and interior oftestis and its duct. . . SAQ 6 Correctthe following sentences if necessary. i) In cyclostomes, sperms develops in seminiferoustubules within the testis. ii) Leydig cells secrete the hormone estrogen. iii) The Inale genital ducts are called fallopian ducts. iv) The accessory sex glands namely prostrate, Cowper's gland, Bulboutheral gland are found in female vertebrates. v) The intromittentorgan in lizard is called gonopodium. tlrinogenital System
- 72. Functional A ~ ~ a t o ~ n y of Chordates - I1 ,411ovarian Ibllicle consislsof an oocyt or immatureegg enclosed in a epithelium. The cells ol~epitheliu~n &re , referredto variously as follicular cell or nurse ccll or granulose cells. In cyclostomes. teleosts and amphibians the epithelium is one layer thick. In the hagfishand in those vertehrates like elasmohranches. reptiles. birds and monotremes in which the oocytcs have heavy yolk deposit. the epithelium appears to be two cells thick. In mammals above thc level nf oionotremes. the tollicular epithelium appears to be many ccll thick. The genital system in vertebrate female consist of: I. Paired gonads -ovaries (sometimes one due to degeneration) 2. Paired oviducts 3. Single female genital opening (Refer to Fig. 9.23 a and b) 9.11.1 Ovary Ovaries lie within the body cavity and are suspended by the dorsal mesentry, the mesovarium, through which pass blood and lytnph vessels and nerves. Primitive vertebrate ovaries are found in hagfish, in which a mesentry-like fold of gonadal tissue stretches across nearly the entire length of the body cavity. In the hagfish, the unique feature is that the fuhctional ovarian tissue occupies only the forward half of the-gonadal mass while the rear part contains the rudimentary testicular tissue. In most fishes except in very primitive forms, the ovaries are similarly elongated. In tetrapods except for mammals the ovaries are usually confined to the middle third or h6lf of the body cavity. particularly during the non breeding seasons. The ovaries of mammals undergl. moderately caudal displacement in order to be located between the kidney and the pelvis (Fig. 9.23 a). The shape of an ovary depends on many factors like whether one egg or thousands are discharged (ovulated), whether the eggs are immature or ripe; whether mature eggs are small or large, or whether pigments occur in the egg cytoplasm, such as those responsible for yellow yolk. The appearance of the ovary is also affected by other factors such as the season of the year in seasonal breeders. (as ovary enlarges during the breeding seasons and diminishes tn size between seasons) the age of the animal (whether juvenile or reproductively active or senile especially in birds and niammals) and the fate of the ovulated egg follicles or sacs. Fig. 9.23: Mammalian female reproductive s)'stcni. (a) saggital view of pelvis of human fcmales 'showing female reproduktive system. (b) cut away of ovary showing internal structures. Ovaries are characterised as saccular or hollow or lacunate (i.e. compartmented) or cotlipact structures. However, most ovaries are similar in construction. They are covered with a germinal epitheliumthat is'continuous with the peritoneum lining the body cavity. Beneath the epithelium the ovary has a layer of connective tissue called tunica albuginea which is considerably thinner than that surrounding the testis. Below this the ovary has an external cortex and internal medulla. The cortex which is the thick outer layer lies immediately internal to tunica albuginea and contains future eggs and at one time or another, eggs in ovarian follicles (developing eggs) (Fig. 9.23 b). The cortex
- 73. also contains remnants of ovulated follicles and in mammals. and clusters of interstitial cells that in some species are glandular. The cortical components are embedded in a supportive frame work of connective, vascular and neural tissue that form the stroma. Internalto the cortex is the vascular, meso-dermalmedulla which consists of blood and lymph vessels, nerves and connective tissue. 'The medulla lacks germinal elements and exhibitsno significant cyclical activity. It is usually inconspicuousand is continuous with the dorsal mesentry. In the cyclostomes the medullaand dorsal mesentry are indistinguishable from each other. On the contrary in mammals the medulla is almost completely surrounded by the cortex and converges on the mesovarium at a narrow hilus, at which nerves and vessels enter the ovary. In the medulla of the mammalian ovary near the hilus arc small masses of blind tubules or solid cords called the rete ovari. The 'rete ovari' are homologous(of the same embryonic origin) with rete testis in the male. The rudimentaryright ovary of the birds usually consists of only medullary tissue. 9.11.2 Female Genital Ducts Eggs are released into the coelo~nic cavity. Once in the coelom, the mature eggs enter the female genital duct called oviduct which parallelsthe archinephricduct in its embryonic course. The oviduct usuallyjoins the urinogenital sinus near the cloaca. Oviducts except in teleosts and some fishes are modifications of the mullerian ducts and develop in every male or female embryo (except in cyclostomes) as a pair of longitudinal ducts. In males, the Mullerian duct disappears or becomes rudimentary. In females, it grows larger to become the reproductive tract or gonoduct. Itssmooth musclesand cilia of the ciliated cells, in its lining propel eggs along the tract. In most teleost fishes with tubular ovaries, eggs do not enter the coelom at large but go directly into a special genital duct called gonaduct which is named so, because it is apparently derived directly from the gonad. The gonaductencloses a small part of the main coelom (Fig. 9.24). In higher mammalsthe oviduct differentiates into three regions (i) Fallopiantube (ii) uterus and (iii) vagina. The oviduct of most other vertebrates is open at both ends, at the urinogenitalsinus which may open into the cloaca and near the ovary by means of infundibulum which is also called pre ampulla. The end next to the ovary is ringedwith mobile, finger like ciliated projections called fimbriae that actively envelopsthe ovary near the time of ovulation (see Fig. 9.24). Oviducts may be long or short and may have glandular portions that are modified to be secretory in function. As a result along the course, the oviducts may differentiate into (i) a region that provide protective and nutrient ~naterial on the eggs, (ii) uterus (pl: uteri) to lodge the developingembryo in case of viviparousanimals. (iii) the terminal segment of the female genital duct which is modified to receive the intromittent organ. Externalcervical os Fig. 9.24: Thc rcproductivc system of human female showing oviduct. ovary, uterus and vagina.
- 74. Ful~ctiona!I: I ! I J , : I ~ .,f' Chordates -11 Uterus is the muscular middle part of the oviduct (Refer to Fig. 9.23 a and Fig. 9.24 again). Its muscles form the myometrium and its inner lining is called endometrium. The endometrium becomes highly vascularised before the blastocyst stage (or developing embryo) implants. In all mammals the uterus narrows to form the posterior cervix (Fig. 9.23 and Fig. 9.24). Lips of the cervix enclosethe uteri opening or '0s uteri' through which sperms rise upto the upper part of oviduct for fertilisation.The cervix dilates during delivery of the baby. Uterus shows a num~ber of variations in mammals. In primitive mammals like monotremes (egg laying mammals) and marsupialsthere are two uteri (duplex uterus). In most mammals hbwever, the distal parts of the two uteri are fused together to give a bipartite or bicornuate uterus. In higher primates there is a complete fusion of the two uteri to form a single simplex uterus (Fig. 9.25). Duplex Uterus - Bipartiteuterus Bicomuateutcrnus Simplex uterus Fig. 9.25: Diagram showing the fusion of posterior ends of paired uteri in females of placental n~ammals. (The uterusAnd part of the vagina have been cut open.) Vagina 'The uterus leads through the narrow cervix into the vagina. Vagina is the fused terminal part of the Mullarian duct. It opens into urinogenitalsinus (Fig. 9.22 ) or is extended to open directly to theoutside. It is a muscular distensibletube which receives the penis during mating. It has convolutions on it which are called vaginal rugae. Vagina opens to the exterior by the female genital opening. Vagina is absent in egg laying mammals. Marsupials,have paired vagina that opens into urinogenital sinus. To match the two vaginae, male penis is forked at the tip and one tip enters one lateral vaginal canal to discharge semen. 9.11.3 Female Accessory Glands You have already read in subsection 9.10.3 about the various accessory glands associated with the male genital system. You will similarly find that the accessory glands are also' associated with the female genital system of many vertebrates.Oviducts of many female vertebrates havcglands in the oviduct called the albumin gland that coat the egg with albumin. Other glands associatea with the oviduct are the shell gland or nidamental gland that cover the egg with shell material; mucous cell or oviducal tubular glands that secrete ajelly like material. Amniotesthat lay large eggs may have in the oviduct mucous secretingglands cal~led vaginal mucous gland that coat the egg prior to expulsion, possibly to lubricate it. Some fishes have adhesive glands that coat the eggs with a sticky secretion so that the egg scan adheretogether or to appropriate objects. In some vertebrates that retain the developing embryos within the body, special glands of the oviduct evolve in order to nourish the young. These glands secrete into the oviduct so that
- 75. young presentthere absorb or ingestthe secretion. This secretion is called uterine milk or en1bryotrophe. 9.11.4 External Female Genitalia In females the external genitalia in comparison to males is feebly developed. Maximum developme~it of female genitalia is in tlie Order Primatesof mammals. In liu~nan females it is well developed and is described below: External Genitalia of Human Female The external genital organs of the female aretermed vulva (Refer to Fig. 9.24).The internalgenitaliaof the female consistsof an outermost structure called labia majora which are a pair of skin folds and contain adiposetissue. Within the cleft formed by these folds are the labia minora, which are a smaller pair of skin folds that are highly vascularised but have no fatty tissue. At the anterior end of the vulva, these two interior skin folds partly enclose tlie clitoris, a small organ for sexual stimulation.The opening of the urethra is about midway betweenthe clitoris and vaginal opening. The vaginal opening is located behind the urinary meatuesand is much larger than the urinary opening. It is covered by a thin mucous membranethe hymen. Bartholin's glands (or bulbovestibular glands) or greater vestibular glands are two bean shaped glands, one on either side ofthe vaginal opening. These secrete a lubricating fluid. The two glands open by a single duct between hymen and labia minora. Bartholin glands are homologousto male bulbourethral glands. A group of tiny mucousglands, the lesser vestibular glands also called Skenes glands open into the vestibule.. 9.11.5 Mammary Apparatus Mammalsare named so because of the characteristic presenceof the mammaryapparatus, which includes(i) mammary glands, (ii) elevated nipples which are the outlets for secretion of these glands and (iii) breasts or mammaewhich are the integumentary .swellingsdue to localised presenceofthese glands (Fig. 9.26). Mammary glands are modified sweat glands. They are present in both male and female mammalsbut they become well developedonly in females as their development is controlled during puberty by the ovarian hormones estrogenand progesterone. Each mammary gland is divided into anumber of lobes and each lobe has several lobules. Lobulesare formed of connectivetissue in which secretory cells called alveoli are embedded. Alveoli are arranged in grape like clusters around minute ducts. Progesteronestimulates the growth of alveoli and estrogenstimulates growth of ducts. ,Nipple Fig. 9.26: Human female hreast showing mammary apparatus. Ducts from lobules units to fonn a single lactiferousor milk carrying duct per lobe. Each duct opens by a pore at the nipple. The nipple is bordered by a pigmentedarea called areola containingseveral sebaceousglands wh,ich appear as small nodules underthe skin. The nipples vary in number and location'in different mammals, The number
- 76. Functional Anatonly of Cl~ordates - I1 depends on the nunbber of offsprings born in a litter. The position of mammae depends on their availability to the suckling offspring. In monkeys and other primates which are arboreal, the mamlnae are pectoral. Humans, who have descended from arboreal ancestors also have pectoral nipples. In ruminants which suckle their young, while standing, the mammae are elongated and project downwards. In pigs they are arranged on the sides and so the mother lies down to suckle the young. The number of nipples vary Horse, bats, whales and humans have a pair of nipple. 9.12 SURVEY OF GONADSIN VERTEBRATES Jawless vertebrates: In the jawless fishes the gonads begin as paired structures but fuse later in development into a medial gonad. Sperms forni in the ampullae rather than in seminiferous tubulbs. In both male and female forms, special genital ducts are absent. The gametes that are reileased from the gonads pass through the coelomic cavity and out of the body via genital pores. The lack of genital ducts appears to be a primitive vertebrate feature. The complete separation of the anus from the urinogenital.tube appears to be a specialisation. Jawed vertebrates Cartilaginous fish -Gonad structure, origin, as well as pattern of genital ducts of .. cartilaginous fishes are typical forjawed vertebrates in general (Fig. 9.27 a and b) and indicate a common origin of vertebrate genital ducts from archinephric tubules. The Mullerian duct originates as a new structure in jawed vertebrates. The testes are of primitive arnpullaty type. Modifications for internal fertilisation include pelvic fins called claspers (Refer subsection 9.10.4). The many variations on internal development among cartilaginous fishes indicate no common pattern or clearly defined relationship to other fishes or to tetrapods. Fig. 9.27: Reproductive system of cartilaginons fishes (a) male urinogeoital system of a shark (b)female reproduttive system of shark Squabrs. The left ovary has been removed. Bony fishes: Mlost extant teleosts have specialised hollow ovaries and testes with genital ducts called gonoducts that are derived from the gonads (Fig. 9.28 a and b). However some bony fishes similar tojawless fish have coelomic transport. In them. extensions of the gonads in the adult produce hollow testes or ovaries that continue as tubes to transport the egg or sperm respectively to exit from pores through the posterior body wall. The testes may be (i) ampullary (or acinar) as in jawless fish and sharks or (ii) tubular as in most tetrapods. Tubular testes and the genital ducts associated with the kidneys seem to be primitive features of bony fishes.
- 77. Kidney Testis Sperm duct Archinephric duct Urinary bladder Urogenital papilla Fig. 9.28: Reproductive systel~~ of bony fial~cs (a) 111aleuri~~ogcl~ital systenls of tnale sea Il~)rsc, Hippocnntp~rs (b) firmale reproductive system of holly fist1Anrict. Amphibians: In amphibians the gonads and urinogenital ducts are basically like those of primitive bony fishes and tetrapods (Fig. 9.29). Some oviparous and all viviparous forms have developed internal fertilisation which may have originally been an adaptation for a terrestrial mode of life. Caecilians appear to be the most of terrestrial of the amphibians in terms of reproductive adaptation. They all have internal fertilisation and appear to have developed mechanisms to protect the egg, by brooding, egg retention or more colnplete viviparity. Living viviparous amphibians include several species of salamander, approximately five species of frogs and about 20 species of caecilians. The oviducts of females of some viviparous species of frogs, salamanders and caecilian produce nutrient secretions called uterine milk for the young. No placenta is however known to develop within the oviduct. Bidder's d Fat bodv ''u Cloaca Fig. 9.29: ReyroQuctive s y s t e ~ ~ ~ of a~opl~ibians (a) male urisogeaital orgalls ol' load Blrfo anrericctnus, showiag bidder's calla1 (b) female uril~ogenital system of toad &fi, with right ovary removed. Reptiles: In reptiles the ~nesonephric tubules are taken over entirely by the testes for sperm transport (Fig. 9.30 a and b). Urine is then carried across by a new duct, the ureter. The oviducts in the females, similar to that of amphibia and most fishes are also not involved with kidney or more appropriately excretory function. The intromittent organs, the hemipenis and penis of the reptiles indicate a clear adaptation towards terrestrial mode of life and hence internal fertilisation that has been phylogenetically continued into mammalian descendants of reptiles. In some species the oviducts of female may retain living sperm for a long period of time after copulation, upto four years in the case of some turtles. IJrinogenitaI System
- 78. Functional Allatom) of Chordates - I1 Fig. 9.30: Reproduclive systems of reptiles (a) urinogenital system of the n~ale lizard Culores(b) female urinoge~~ital system of lizard, Calotrs. Birds: Birds typically have reptilian reproductive organs except that in females only the left gonad develops in to an ovary (Fig. 9.27 c). The right gonad develops upto a certain stage, after which it regresses and remains undifferentiated. A penis occurs only in primitive birds. Fig. 9.31: Reproduttive system ofAves (a) male urinogenital system of pigeon, Colrmrba(b) female uri~~ogenlital system of Colrmtba. 'The oviduct at its posterior end has pockets called sperm nests that store sperm. As the egg leaves the ovary, it enters the infundibulum where it is fertilised. The fertilised egg is forced along the oviduct in which a spiral band of material called chalazae is added at each end, of the egg, as are also added thick and thin layers of albumen, shell membranes and the shell, around the egg. Mammals Primitive mammals like the rnonotrernes(egg laying mammals) generally have a reptilian reproductive apparatus except for the fact that the penis is tubular instead of being grooved and the lnalnmqry glands are present. Monotreme have a distinct cloaca. The ureter in lnonotremes similar to reptiles and birds lies between the Wolffian or Mullerian ducts instead of lying lateral to the reproductive duct as in eutherian mammals. The oviducts of monotremes are relatively unspecialised as compared to most mammals. They are unfused and open individually into the urinogenital sinus. The oviducts are specialised to the extent that they produfe a shell similar to that of reptilian eggs and also secrete uterine milk called embryotirophe which is absorbed by the embryo and used for nutrition.
- 79. The details of the reproductive system of the rest (Fig. 9.32 a and b) of the mammal groups have been given in various sections of this unit, however a figure of the reproductive systern of a typical mam~iialis given below. Kidney Ureter ' Ampullary gland Sem~nal vesicle Coagulat~nggland Prostategland Col/per's gland Urinary bladder Preputial gland Vas dererens - Glans 4 Testis Anus - Fig. 9.32: Reproductive 3ystem of'rnatnnlal(a) tnale ~~l-itlogential systern ot't~tale rat. Rt~llrrs (b) l'c~~~alc urinogrnitsl system of female rat Rollus. SAQ 7 Fill in the blanks and colnpare your answers with those given at the end of this unit. .................. ........................ i) The four types of ~na~nmalian uteri are ....., ...., .............................and .......................... ii) The muscle layer of the uterus is called ............................... iii) The sequence of organs of mammalian female genital system are: two ovaries - 3 .......................- > .................... ..................- > genital opening. + ........................ iv) Caecilians among amphibians have fertilisation. v) The oviducts of females of some viviparous species of amphibian produce nutrient ............................... secretions called for the young. ........................ vi) In female birds only the gonad develops into the ovary. ................................ vii) The oviducts of lnonotreme secrete uterine milk called 9.13 SUMMARY Both the urinary and reproductive organs arise e~nbryologically from the same or adjacent tissue and maintain close anatomical and sometimes functional association, throughout the organise life. Several types of kidneys are found within the chordate groups. The organs of excretion of the protochordates Branchisfonza(Cephalochordata) and Herdnlania (Urochordata) show no relationship to any part of the vertebrate kidney or other know11fluid regulatory structure. The excretory organs of the vertebrates consist of paired kidneys and their associated ducts. The various types of kidneys that are found in the vertebrates have been derived from a primitive structure termed as archinephros or holonephros. The archinephros (holonephros) consisted of paired archinephric ducts which extended the length of the coelom and were joined by segmentally in arranged tubules, one pair to each segment. The free end of each tubule opened into the coelom by means of a ciliated, funnel shaped nephrostome. Each tubule was intimately associated with a small knot of inter arterial capillaries known as glomerulus. The larval stages of the hagfish and calcilians exhibit an archinephric condition.
- 80. Kidneys of living vertebrates have developed from this primitive plan of the archinephros type of kidney. The various types of vertebrate kidneys may be regarded as successive stages that have evolved in a craniocaudal direction from the original archinephros. They may be clearly observed during the embryonic development of the amniote vertebrate where there is a succession of three developmental stages of kidney - pronephros, mesonephros and metanephros. Some but not all these stages are also seen in other vertebrate groups. In adult ana~nniotes - cyclostomes, fishes and amphibians, the anterior part of the primitive archinephros usually becomes modified or degenerates. In the embryo it appears as a transitory structure called the pronephros. In a few lower vertebrates the pronephros persists in the adult stage and is called the head kidney. A head kidney is found in the adult hagfish and in certain teleosts. The part of the anamniote kidney that remains and forms the adult kidney and is called the opisthonephros. ~his'retains the archinephric duct but differs from the pronephros in that several kidney tubules may be present In each segment and the tubules lose their peritoneal connections. In the opisthonephric kidney there is a general tendency for concentration of kidney tubules towardsthe posterior end. The anterior end loses its importance as an excretory organ and in males is the appropriate by the reproductive system, with the result that the archinephric duct becomes known as ductus deferens. In arnniotes. of the three types of kidneys - pronephros, mesonephros and metanephros which appear in craniocaudal direction during embryonic develo ment, only the metanephros persists to form the adult kidney. The archinephric duct n amniotes is termed as Wolffian duct. P In male amniotes, the Wollfian duct gives rise to the epididymis, ductus deferens and certain other parts of the reproductive system. The urinary system of most vertebrates includes two kidneys and two ureters. A urinary bladder and urethra occur in all mammals and some vertebrates. Ducts from the kidney lead to cloaca in most forms. In teleost fishes they open directly to the outside. In marnmals ducts of the kidney enter the urinary bladder (except in monotremes). Urinary bladder, may be present as ventral out pocketings of the wall of the cloaca. In mammals the bladder opens to the outside through a urethra which in males of all forms except far monotermes is also used by the reproductive system. Each mammalian kidney as well as that of most vertebrates is composed of a renal capsule, enclosing a cortex and an internal medulla. Numerous individual tubules called uriniferous tubules are the basic excretory unit of the kidney which produce urine. Each uriniferous tubules consists of i) nephron,an excretory unit ii) collecting tubule. Each nephron consists of a cup shaped Bowman's or renal capsule, proximal convulated tubwle (PCT), loop of Henle and distal convoluted tubule (DCT). Blood vessels associated with the nephron are an afferent arteriole, glomerular capill,aries, an efferent arteriole, and peritubular capillaries. Kidneys are the principal organs of excretion and osmoregulation in vertebrates. The various functions of the urinary system are (i) excretion of metabolic waste products (ii) regulation of fluid, electrolyte balance and blood pressure (iii) sicretion of hormone erythropoetin which is associated with production of blood cells in case of loss of blood or hypoxia. The kidneys of vertebrates are confronted with two kinds of problems since they occupy diverse habitats like those in which water (i) may not be available for example in terrestrial environment or marine environment or (ii) may be in abundance for instance in fresh water environment. Thus the kidneys and tubules of vertebrates are adapted according to the environment they inhabit. The urinogenital systems of various groups also differ to some extent. :NITALSYSTEM The reproductive system of chordates consist of primary sex organs, the gonads which produce gametes. The primary sex organs or gonads are testes in male and ovaries in female. The testes produce the spermatozoa and the ovaries the ova.
- 81. The vertebrate reproductive system consist of primary and accessory sex organs. The accessorysex organsare ducts and glands which provide passage for transport o f gametes (eggs or spermatozoato the outside of the body). The archinephric duct in anamniotes or the Wolffian duct in amniotes becomesthe reproductiveduct or ductus deferens in the male. In females the Mullerian duct forms the oviduct. Ducts are absent in amphioxus and cyclostomes. In males, spermatozoa are formed within seminiferousampullae or tubules in the testes. Each ductus deferens establishes connections with the testes usually by means o f epididy~uis and persistent kidney tubules called efferent ductules. In anamniotes the epididymis and ductus deferens consistso f parts o f the archinephric duct which may serve in some cases as an excretory duct as well as for passageo fthe spermatozoa. In amniotes l~owever the epididynlis and ductus deferensare formed from the persistent Wolffian duct. which is entirely dissociatedfrom the ureter o f tlle adult metanephric kidney. The testes in vertebratesare located in the abdominal cavity, except in a number o f mammals. In most mammals they are eithertemporarily or permanently locatedin a scrotum outsidethe body proper. In male vertebratesespecially mammals accessory glands are associated with primary sex organs. These secrete seminal fluid in which spermatozoaare suspended and which are essential for thc viability of the spermatozoaand for transport through the reproductive tract both male and female. Fertilisatio~i in vertebrates nlay be external or internal. Most vertebrates with internal fertilisation havecopulatory organs which are used in transferring spermatozoa from male to female. In fishes these usually consist o f modified fins. In snakes and lizards pairedhemipenisare present; but in turtles, crocodilians, certain birds and all mammals. a single penis is used for this purpose. The urethra * corning from the urinary bladder passes through the penis in all mammalsexcept in monotremes. It thus serves for passage of both urinary and seminal fluids. Eggs develop within ovaries and when fully developedthey break out ofthe ovaries into the coelo~n. Each oviduct generally opens into tile coelom by afunnel-shaped ostium. The oviducts of teleosts are derived in a different manner and may not be homologous with those o f other vertebrates. In forms below rnammals the pairedoviducts are separate and usually open independently into a cloaca. In most higher mammalsa cloaca is absent in the adult and each oviduct differentiates into three regions (i) fallopian tube (ii) uterus (iii) . vagina. Various degrees of fusion o f the paired uterus occur in mammals, resulting in different types o f uteri. The external genitalia o f females called vulva is feebly developed in co~nparison to males. Maximum development of female genitalia is in the primates. Accessory glands are associatedwith the female reproductiveand secrete mucous. The parallel developmentof male and female reproductive system is very striking. The various structures in one sex have obvious homologouswith those o f the opposite sex. 9.14 TERMINAL QUESTIONS I. Why are the urinary and reproductive systems usually studied under a common urinogenital system? ................................................................................................... llrinogenitnl System
- 82. Functional Anatomy of Chordates - Il 2. Draw a well labelled diagram of the uriniferous tubule of mammal. 3. Label the given diagram of human kidney. 4. Write short notes on (a) kidney blood circulation (b) types of mammalian uteri. (a)...................................................:............................................. ................................................................................................... (b)................................................................................................ ................................................................................................... 5. What are intromittentorgans? Describe the reptilian intromittent organ. ...................................................................................................
- 83. llrinogc~~ital System 6. List the functions of the excretory systems. ................................................................................................... 7. Describe the a~nniote testis. ................................................................................................... ................................................................................................... ................................................................................................... ................................................................................................... ................................................................................................... ...................................................................................................
- 84. Functional Xaatoniy ol' Chordates i l l 9.15 ANSWERS Self-assessmentQuestions 1. i) glomerulus; ii) hilum or hilus; iii) renal; iv) ~nesoderrnal 2. i) a) nephron; b) glomerulus; c) loop of Henle d) duct of Belleni ii) renal corpuscle/glomerulus and Bowman's capsule, proxi~nal convoluted tubule, descending limb, Henle's loop, ascending limb, distal convoluted tubule, collecting tubule. iii) a, b, c iv) A B a) capillaries of efferent arterioles vas a rectae b) renal interstitium prostaglandin c) macula densa juxta glomerular apparatus d) erythropoietin red blood cells 3. a-i; b-i;c-1 4. i) testis ovary; ii) secondary; iii) Wolffian/Mesonepliric; iv) ovaries; v) cloaca; vi) mesorchium; vii) intromitten. 5. i) mesoderm; ii) primary; iii) oviparous; iv) viviparous; v) female 6. i) In cyclostomes sperms develop in seminiferous ampullae of testis. ii) Leydig cells secrete the hormone testosterone. iii) The male genital ducts are called ductus deferens or vas defens. iv) The accessory sex glands, prostrate glands and bulboutheral glands are found in male vertebrates. v) The intromittent organ in lizard is called hemipenes. 7. i) Duplex, bipartite, bicornuate, simplex: ii) Myometrium; iii) Two oviducts (fallopian tubes) - > uterus - > vagina; iv) internal; v) uterine milk; vi) left; vii) embryotrophe Terminal Questions 1. Through the urinrary and reproductive system have nothing in common functionally; they are usually studied under a common urinogenital system because both develop from the some segmental blocks of trunk mesoderm or adjacent tissues and share many of the ducts. 2. Draw figure on basis of Fig. 9.6 of this unit. 3. Label figure on basis of Fig. 9.9. 4. Refer a) subsection 9.5.2; b) subsection 9.11.2 - uterus. 5. Refer subsection 9.10.4. Intromittent organs of amniotes. 6. Refer Fig. 9.14 (b). 7. Refer section 9.10 testis and seminiferous tubules.